

Metabolic Diversity in Human Non-Small Cell Lung Cancer Cells
- PMID: 31564558
- PMCID: PMC6898782
- DOI: 10.1016/j.molcel.2019.08.028
Metabolic Diversity in Human Non-Small Cell Lung Cancer Cells
Abstract
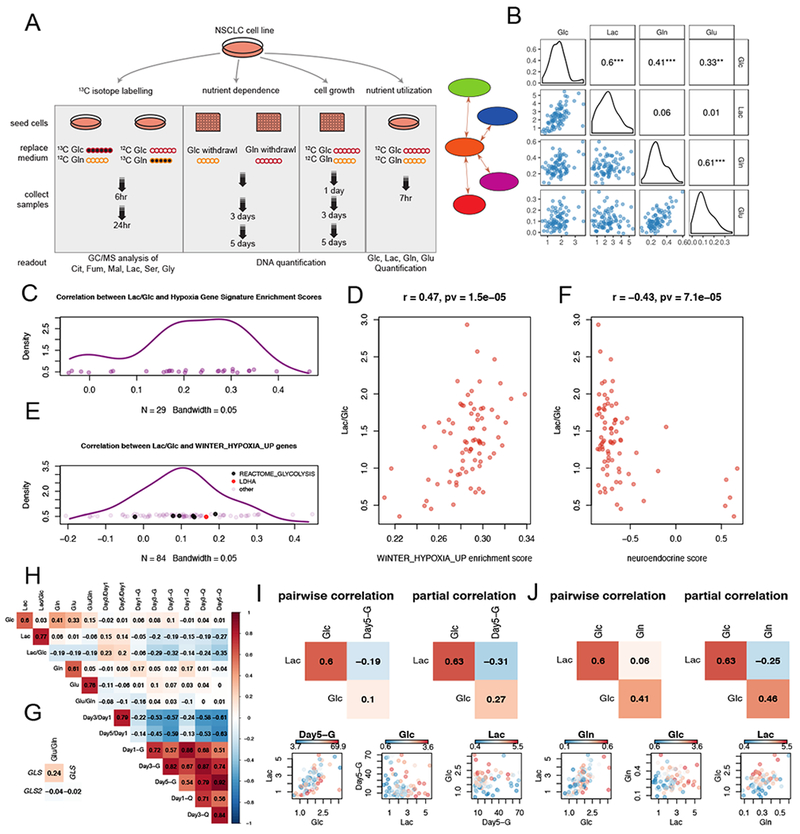
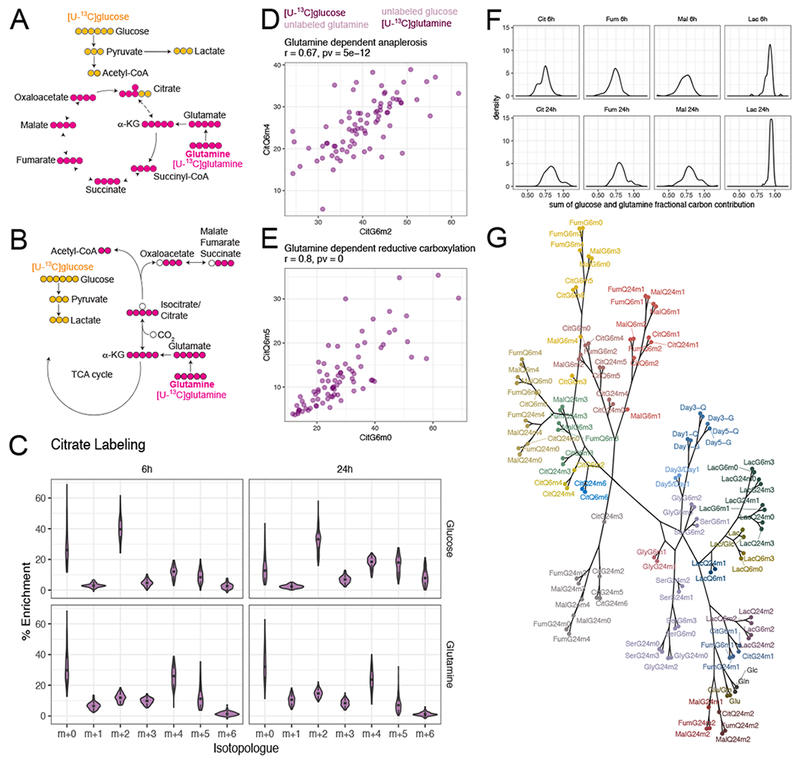
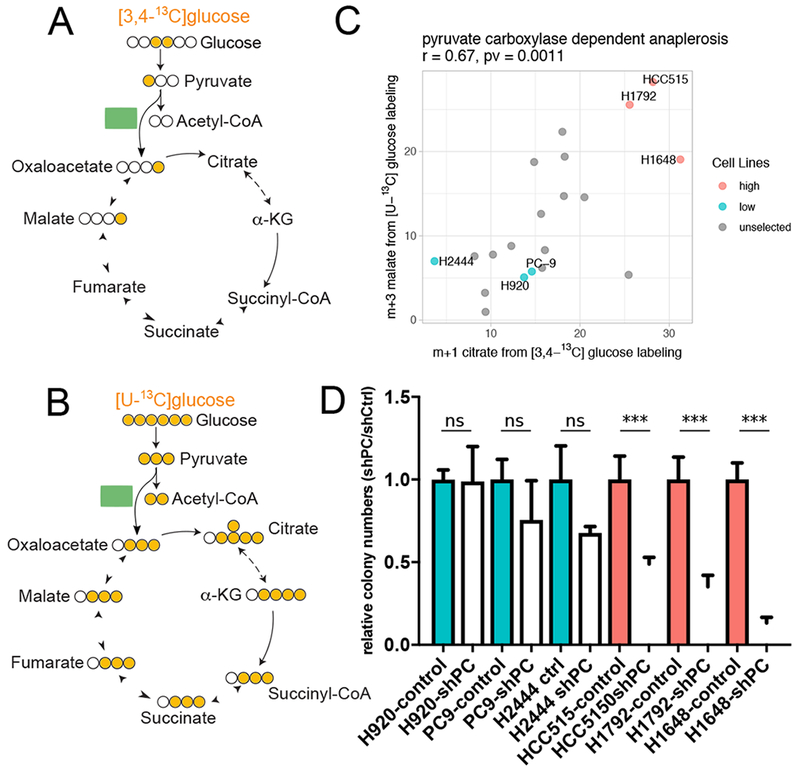
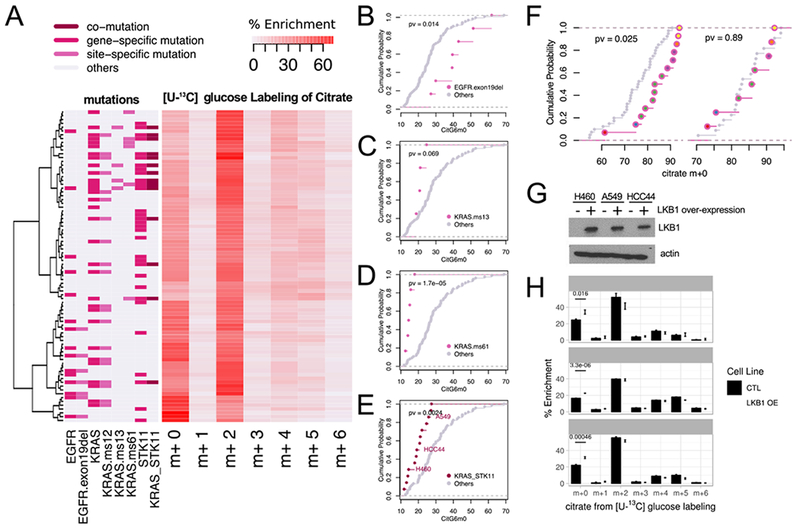
Intermediary metabolism in cancer cells is regulated by diverse cell-autonomous processes, including signal transduction and gene expression patterns, arising from specific oncogenotypes and cell lineages. Although it is well established that metabolic reprogramming is a hallmark of cancer, we lack a full view of the diversity of metabolic programs in cancer cells and an unbiased assessment of the associations between metabolic pathway preferences and other cell-autonomous processes. Here, we quantified metabolic features, mostly from the 13C enrichment of molecules from central carbon metabolism, in over 80 non-small cell lung cancer (NSCLC) cell lines cultured under identical conditions. Because these cell lines were extensively annotated for oncogenotype, gene expression, protein expression, and therapeutic sensitivity, the resulting database enables the user to uncover new relationships between metabolism and these orthogonal processes.
Keywords: (13)C stable isotope labeling; cancer metabolism; cell lines; gene expression; glucose; glutamine; non-small cell lung cancer; oncogenotypes; protein expression; therapeutic sensitivity.
Copyright © 2019 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
LKB1 and KEAP1/NRF2 Pathways Cooperatively Promote Metabolic Reprogramming with Enhanced Glutamine Dependence in KRAS-Mutant Lung Adenocarcinoma.Cancer Res. 2019 Jul 1;79(13):3251-3267. doi: 10.1158/0008-5472.CAN-18-3527. Epub 2019 Apr 30. Cancer Res. 2019. PMID: 31040157 Free PMC article.
-
Metabolic profiling of potential lung cancer biomarkers using bronchoalveolar lavage fluid and the integrated direct infusion/ gas chromatography mass spectrometry platform.J Proteomics. 2016 Aug 11;145:197-206. doi: 10.1016/j.jprot.2016.05.030. Epub 2016 May 30. J Proteomics. 2016. PMID: 27255828
-
Quantitative proteomic approach to understand metabolic adaptation in non-small cell lung cancer.J Proteome Res. 2014 Nov 7;13(11):4695-704. doi: 10.1021/pr500327v. Epub 2014 Aug 25. J Proteome Res. 2014. PMID: 25029028
-
Glutamine reliance in cell metabolism.Exp Mol Med. 2020 Sep;52(9):1496-1516. doi: 10.1038/s12276-020-00504-8. Epub 2020 Sep 17. Exp Mol Med. 2020. PMID: 32943735 Free PMC article. Review.
-
Alterations of metabolic genes and metabolites in cancer.Semin Cell Dev Biol. 2012 Jun;23(4):370-80. doi: 10.1016/j.semcdb.2012.01.013. Epub 2012 Jan 28. Semin Cell Dev Biol. 2012. PMID: 22306135 Free PMC article. Review.
Cited by
-
Construction of the metabolism-related models for predicting prognosis and infiltrating immune phenotype in lung squamous cell carcinoma.J Cancer. 2023 Oct 24;14(18):3539-3549. doi: 10.7150/jca.86942. eCollection 2023. J Cancer. 2023. PMID: 38021151 Free PMC article.
-
Lung Cancer Computational Biology and Resources.Cold Spring Harb Perspect Med. 2022 Feb 1;12(2):a038273. doi: 10.1101/cshperspect.a038273. Cold Spring Harb Perspect Med. 2022. PMID: 34751162 Free PMC article. Review.
-
Insight of a Metabolic Prognostic Model to Identify Tumor Environment and Drug Vulnerability for Lung Adenocarcinoma.Front Immunol. 2022 Jun 23;13:872910. doi: 10.3389/fimmu.2022.872910. eCollection 2022. Front Immunol. 2022. PMID: 35812404 Free PMC article.
-
Long Noncoding RNAs and Circular RNAs in the Metabolic Reprogramming of Lung Cancer: Functions, Mechanisms, and Clinical Potential.Oxid Med Cell Longev. 2022 Jun 15;2022:4802338. doi: 10.1155/2022/4802338. eCollection 2022. Oxid Med Cell Longev. 2022. PMID: 35757505 Free PMC article. Review.
-
miR-183-5p regulates ECM and EMT to promote non-small cell lung cancer progression by targeting LOXL4.J Thorac Dis. 2023 Apr 28;15(4):1734-1748. doi: 10.21037/jtd-23-329. Epub 2023 Apr 7. J Thorac Dis. 2023. PMID: 37197500 Free PMC article.
References
-
- Byers LA, Diao L, Wang J, Saintigny P, Girard L, Peyton M, Shen L, Fan Y, Giri U, Tumula PK, et al. (2013). An epithelial-mesenchymal transition gene signature predicts resistance to EGFR and PI3K inhibitors and identifies Axl as a therapeutic target for overcoming EGFR inhibitor resistance. Clin Cancer Res 19, 279–290. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
