

Resveratrol Improves Boar Sperm Quality via 5'AMP-Activated Protein Kinase Activation during Cryopreservation
- PMID: 31565152
- PMCID: PMC6746164
- DOI: 10.1155/2019/5921503
Resveratrol Improves Boar Sperm Quality via 5'AMP-Activated Protein Kinase Activation during Cryopreservation
Abstract
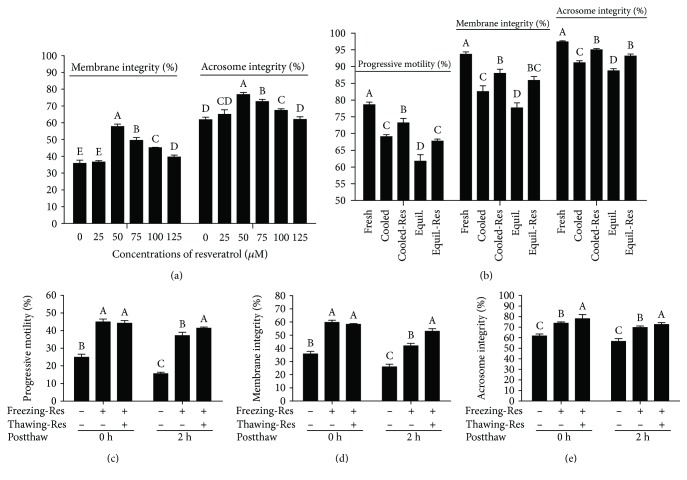
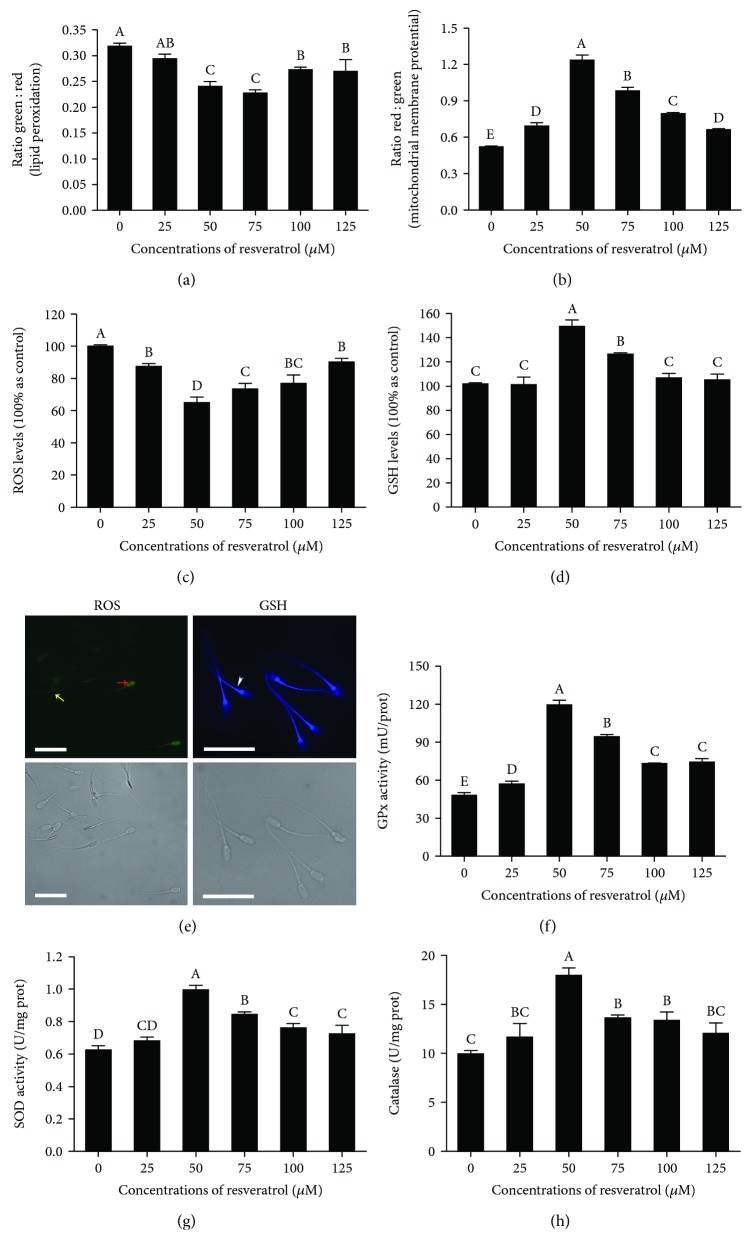
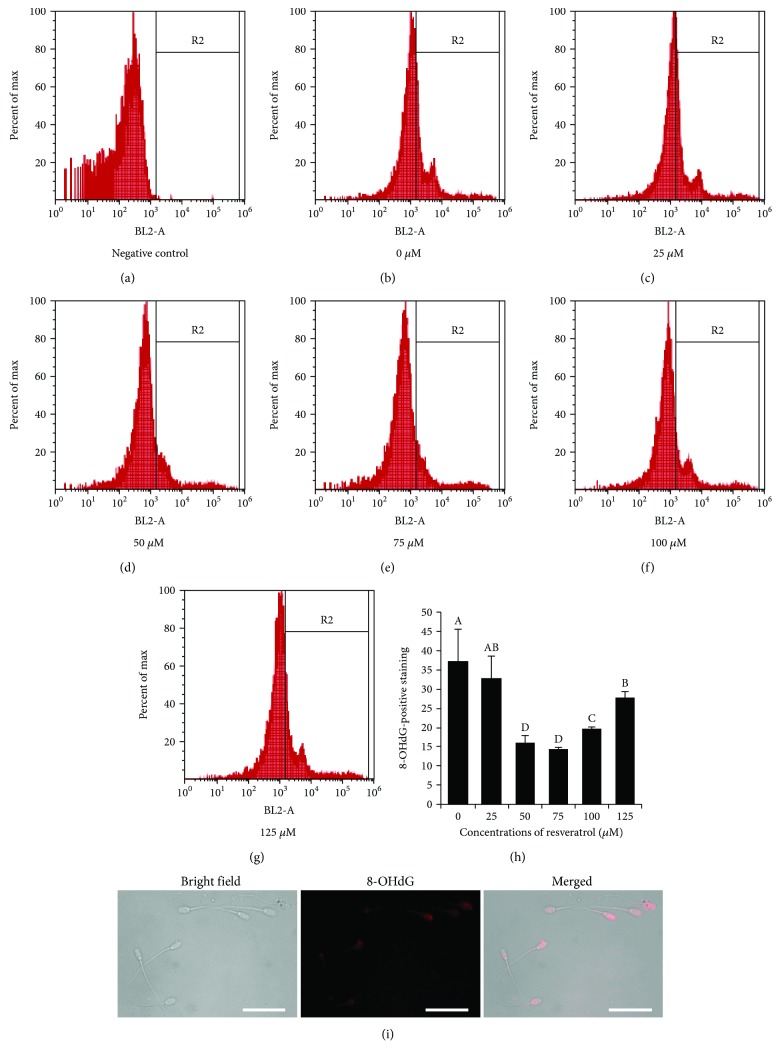
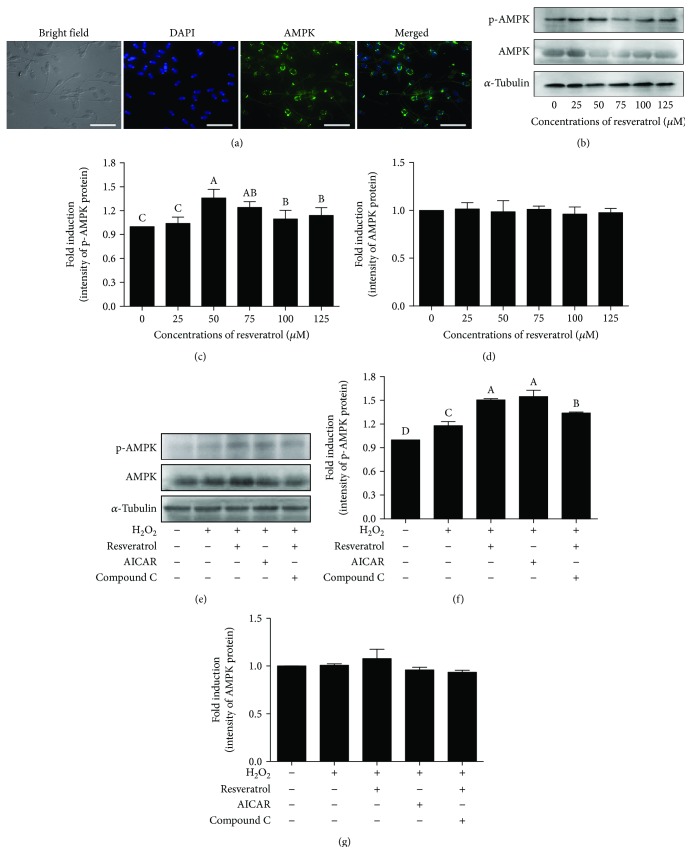
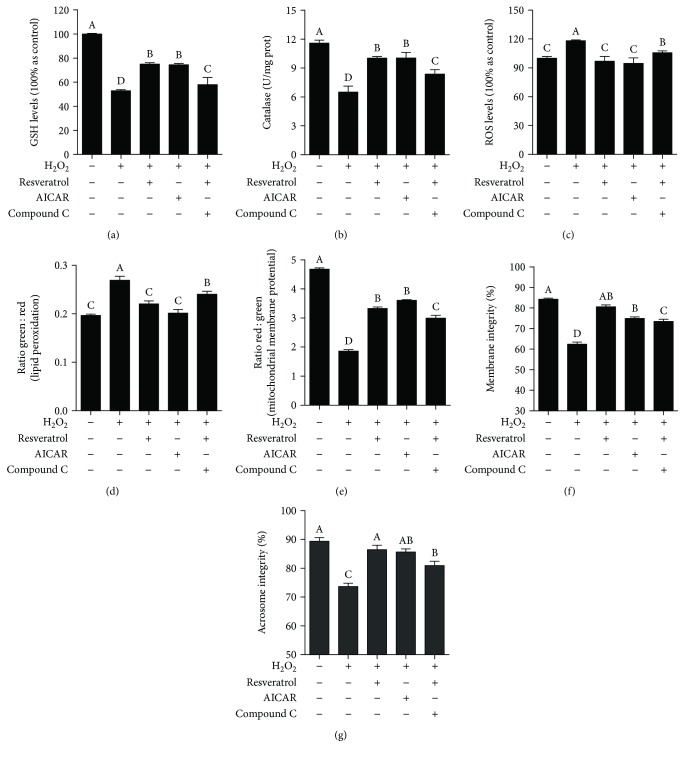
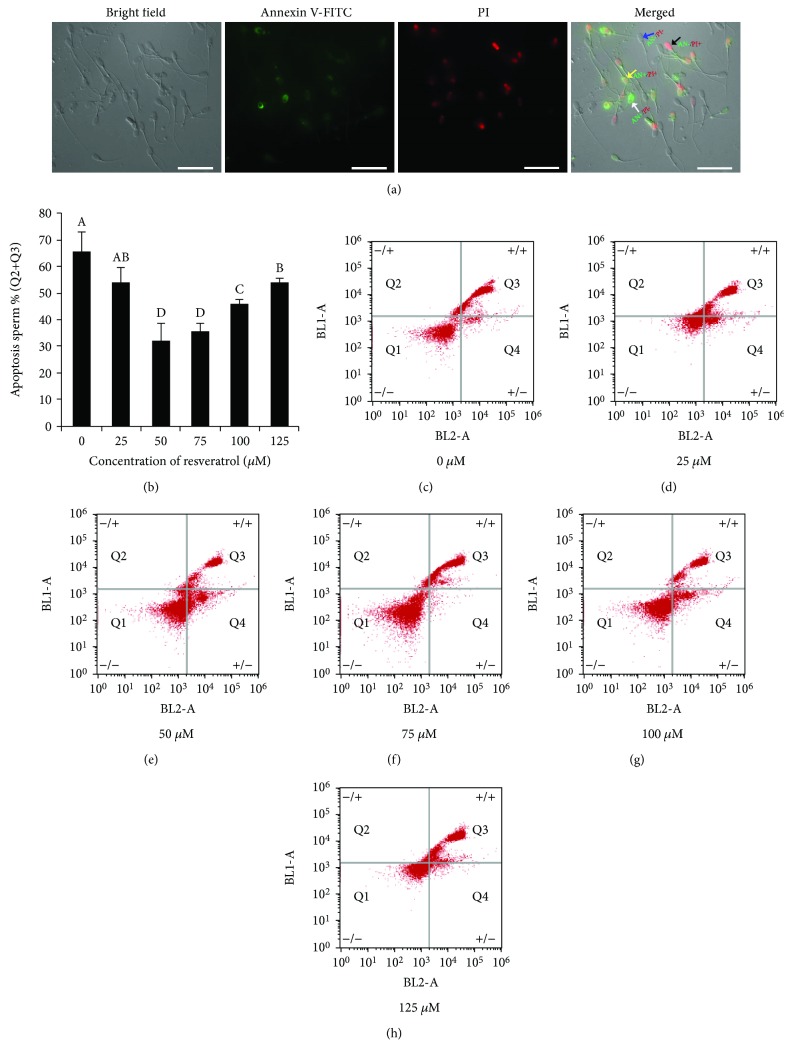
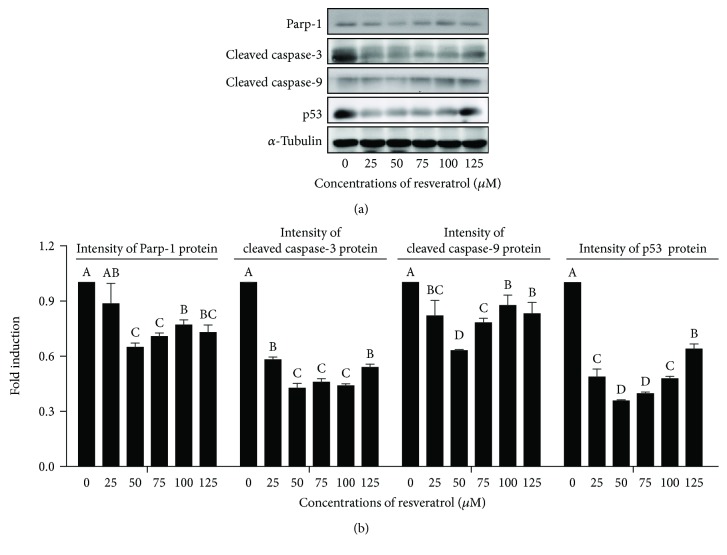
Mammalian sperm is highly susceptible to the reactive oxygen species (ROS) stress caused by biochemical and physical modifications during the cryopreservation process. 5'AMP-activated protein kinase (AMPK) is involved in regulating both cell metabolism and cellular redox status. The aim of the present study was to investigate whether the resveratrol protects boar sperm against ROS stress via activation of AMPK during cryopreservation. Boar sperm was diluted with the freezing medium supplemented with resveratrol at different concentrations (0, 25, 50, 75, 100, and 125 μM). It was observed that the addition of 50 μM resveratrol significantly improved the postthaw sperm progressive motility, membrane integrity, acrosome integrity, mitochondrial activity, glutathione (GSH) level, activities of enzymatic antioxidants (glutathione peroxidase (GPx), superoxide dismutase (SOD), and catalase), and the phosphorylation of AMPK. Meanwhile, the lipid peroxidation, ROS levels, and apoptosis of postthaw sperm were reduced in the presence of 50 μM resveratrol. Furthermore, when fresh boar sperm was incubated with the medium in the presence of 50 μM resveratrol and 30 μM Compound C (an AMPK inhibitor), the effects of the resveratrol were partly counteracted by the Compound C. These observations suggest that the resveratrol protects boar sperm via promoting AMPK phosphorylation. In conclusion, the addition of resveratrol to the freezing extenders protects boar sperm against ROS damage via promoting AMPK phosphorylation for decreasing the ROS production and improving the antioxidative defense system of postthaw sperm. These findings provide novel insights into understanding the mechanisms of resveratrol on how to protect boar sperm quality contrary to the ROS production during cryopreservation.
Copyright © 2019 Zhendong Zhu et al.
Conflict of interest statement
The authors have nothing to disclose.
Figures
References
-
- Severo C. K., Pereira G. R., Pereira A. M., et al. Cysteine addition on short-term cooled boar semen preservation and its relationship with swine field fertility. Pesquisa Veterinária Brasileira. 2011;31(suppl 1):25–32. doi: 10.1590/S0100-736X2011001300005. - DOI
-
- Mazzilli F., Rossi T., Sabatini L., et al. Human sperm cryopreservation and reactive oxygen species (ROS) production. Acta Europaea Fertilitatis. 1995;26(4):145–148. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
