

O-GlcNAcylation and phosphorylation of β-actin Ser199 in diabetic nephropathy
- PMID: 31566433
- PMCID: PMC6879942
- DOI: 10.1152/ajprenal.00566.2018
O-GlcNAcylation and phosphorylation of β-actin Ser199 in diabetic nephropathy
Abstract
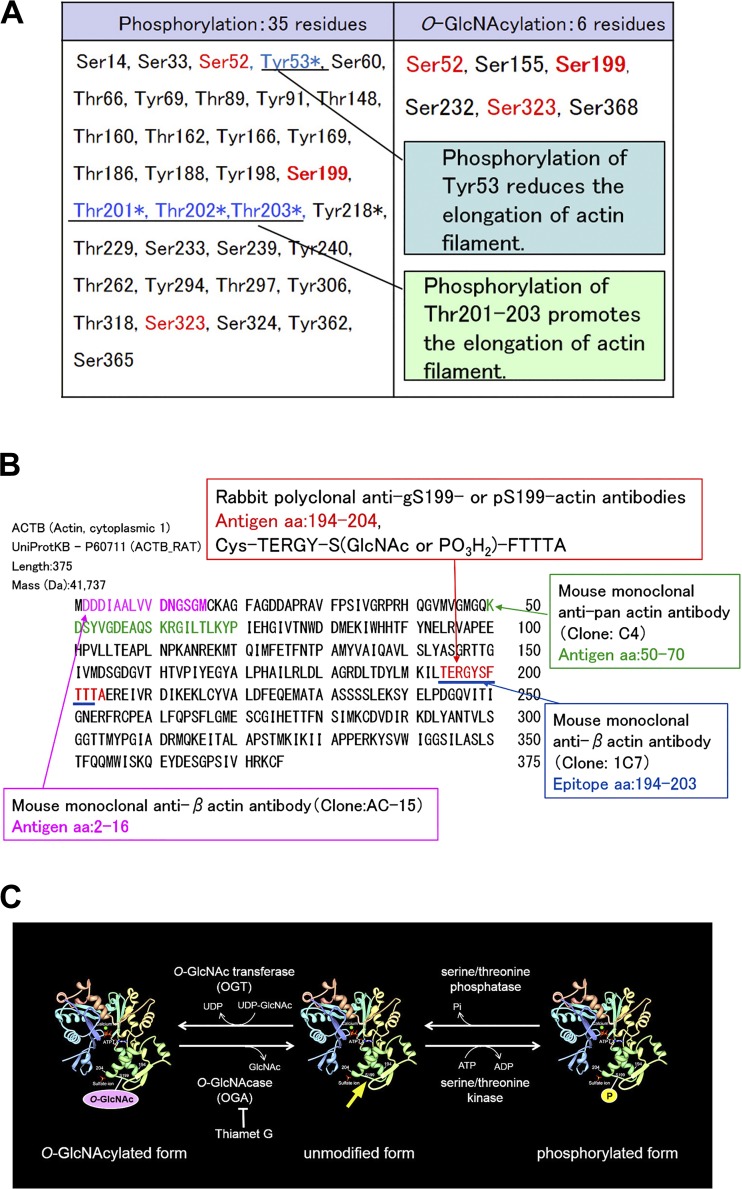
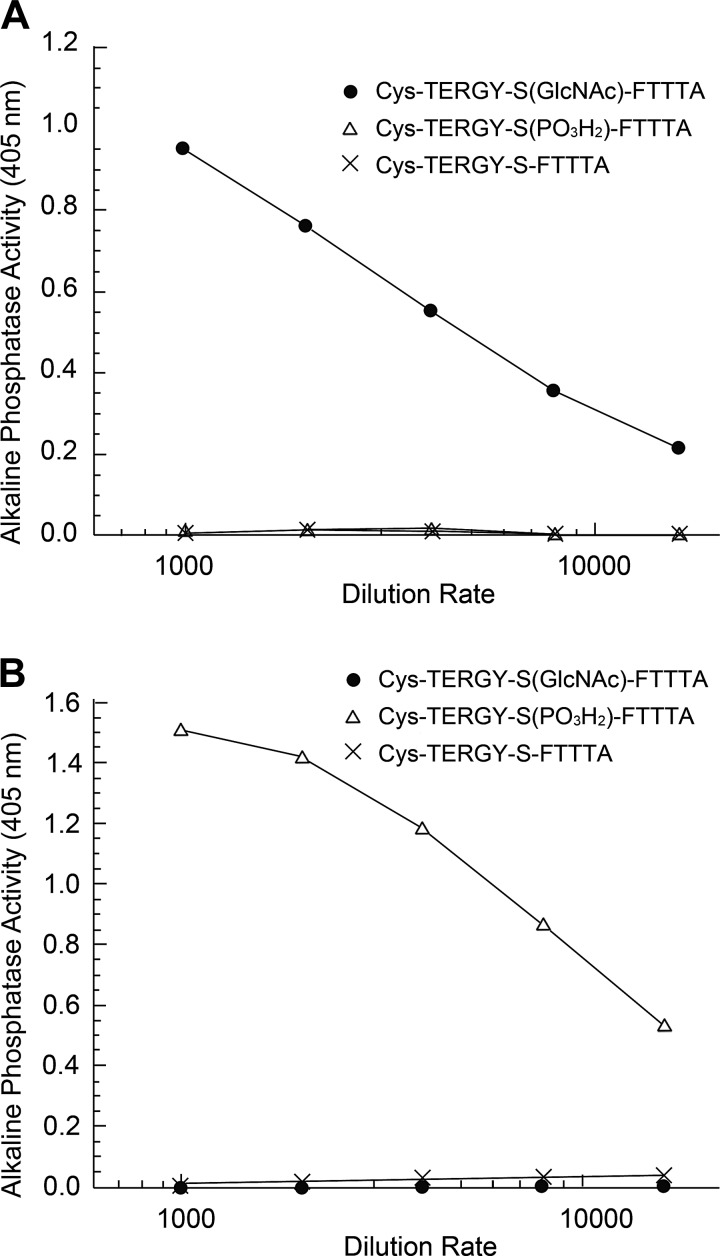
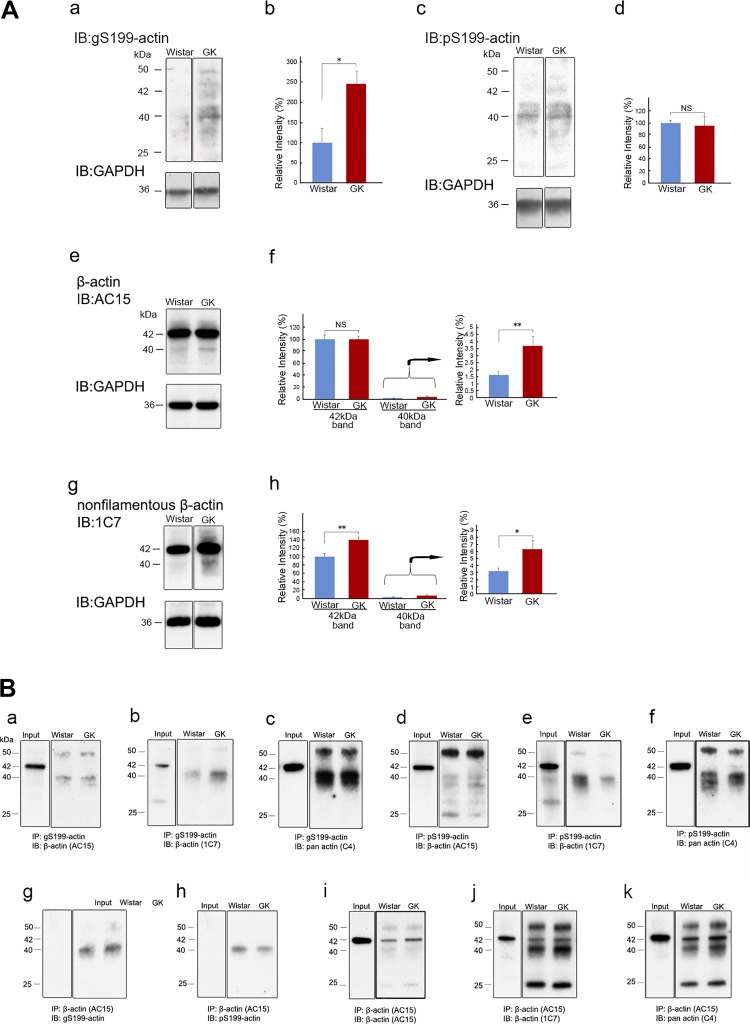
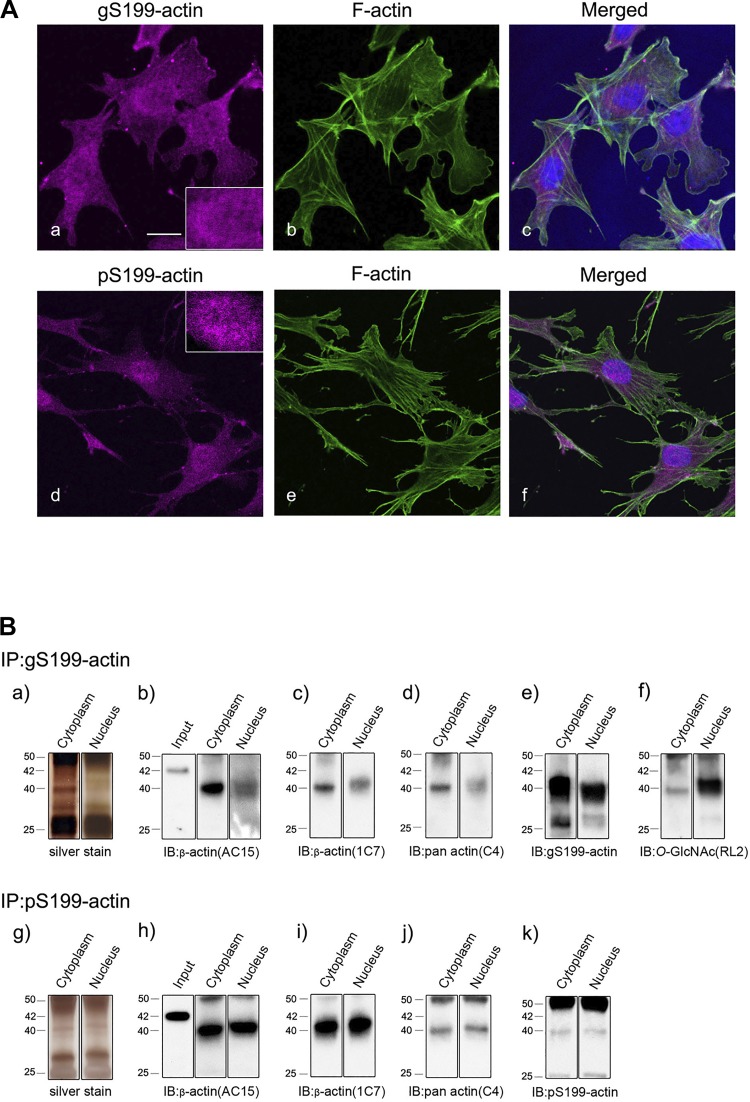
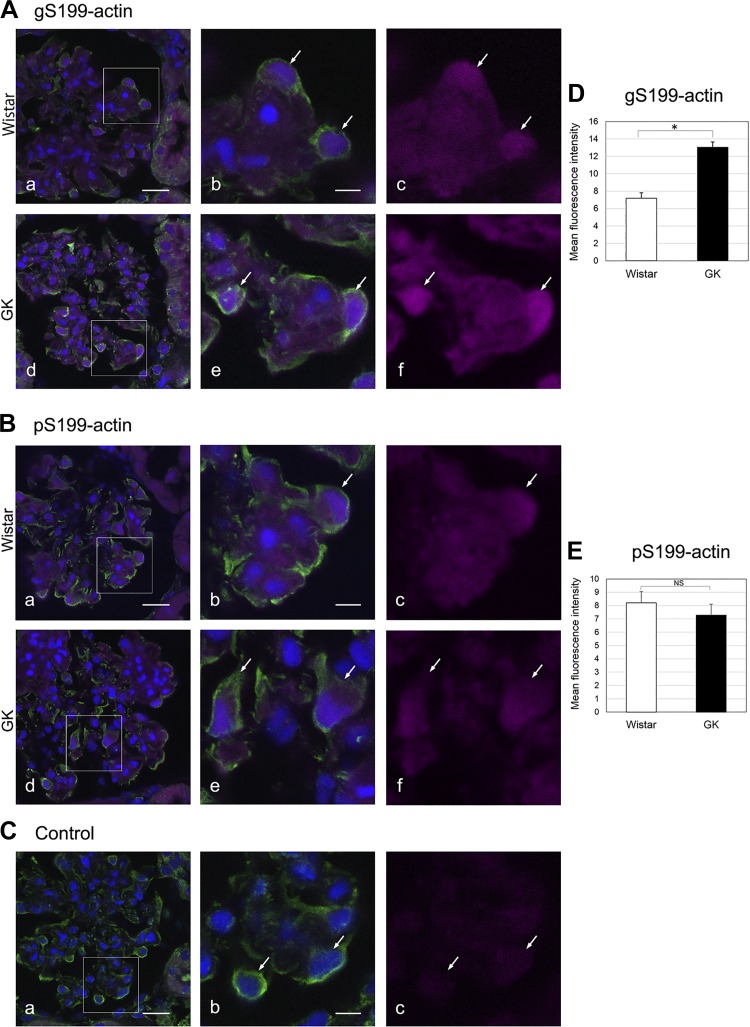
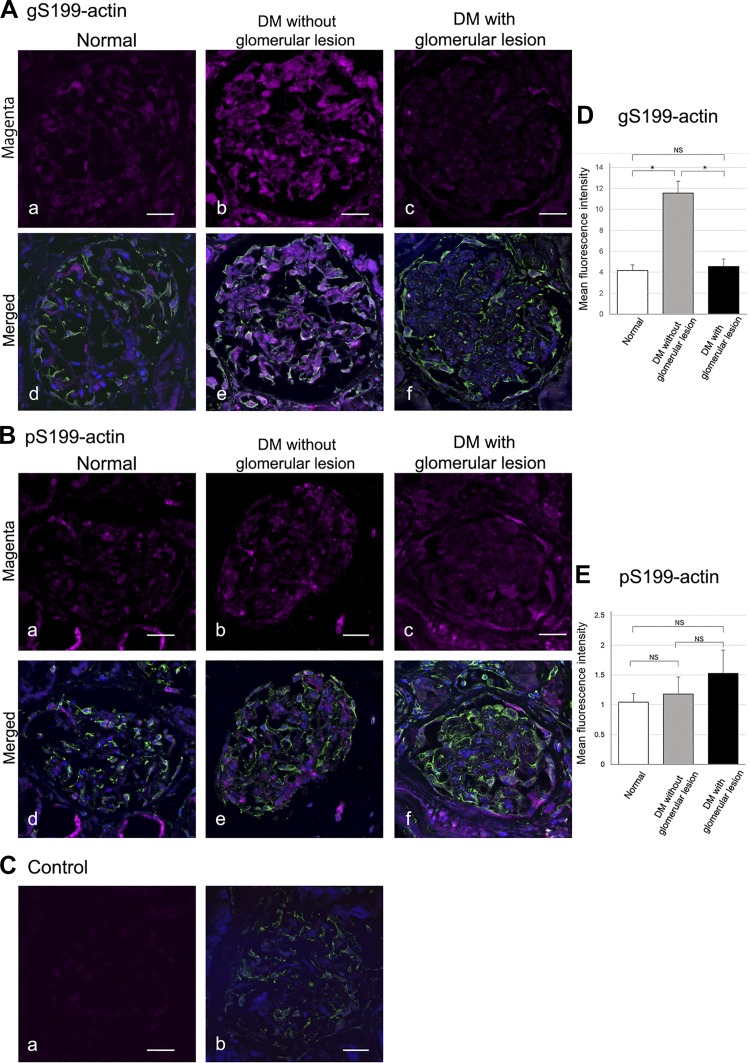
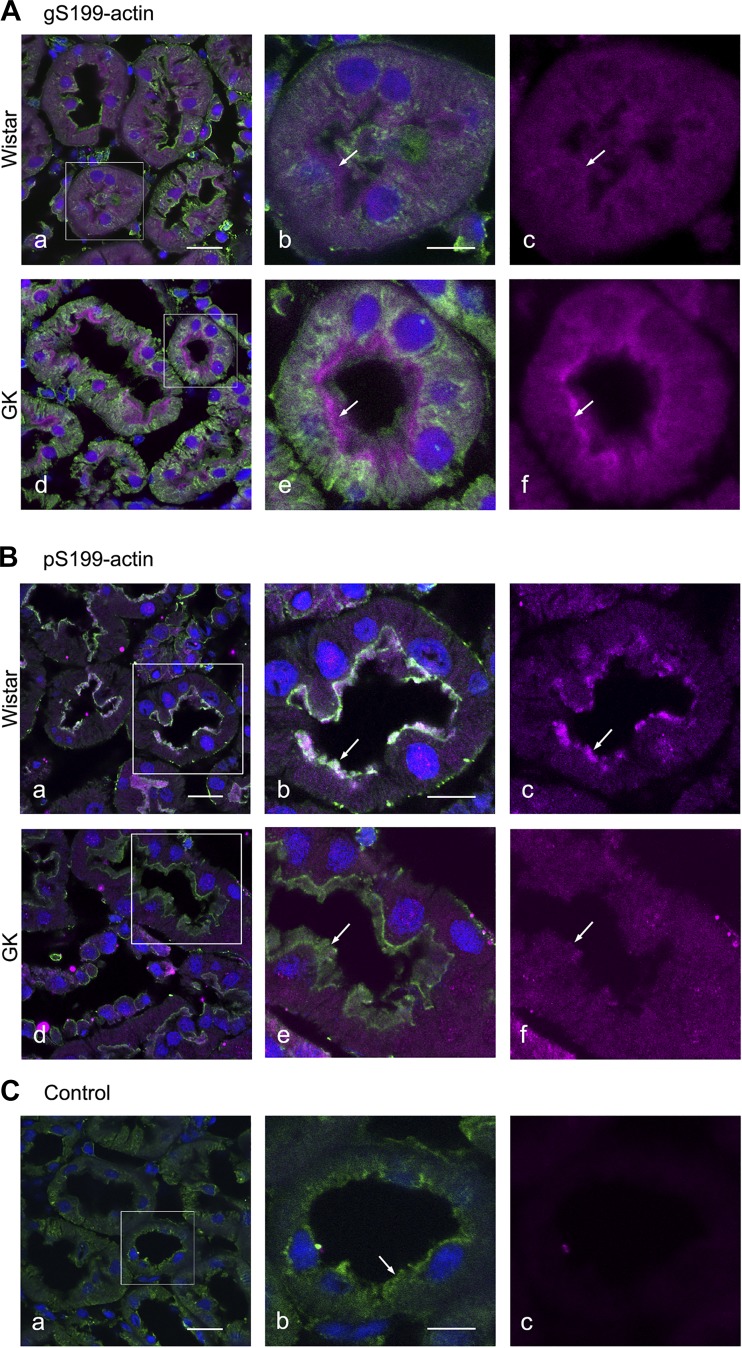
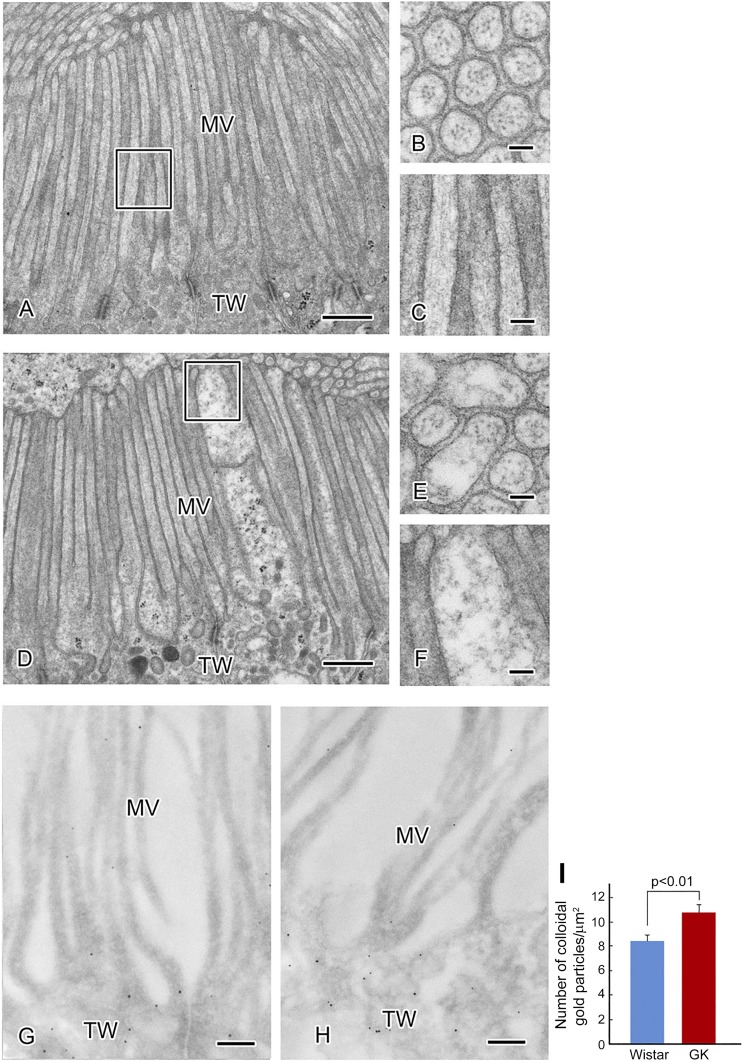
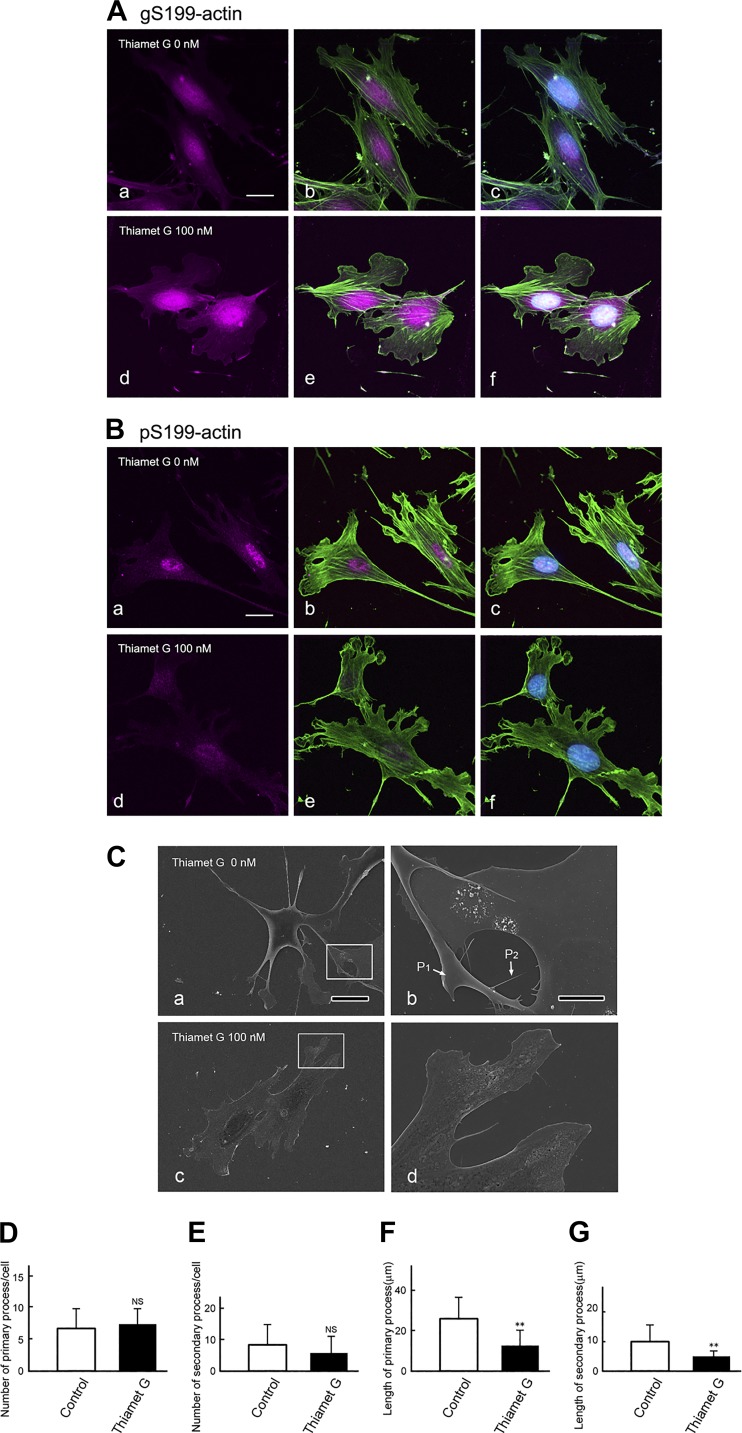
The function of actin is regulated by various posttranslational modifications. We have previously shown that in the kidneys of nonobese type 2 diabetes model Goto-Kakizaki rats, increased O-GlcNAcylation of β-actin protein is observed. It has also been reported that both O-GlcNAcylation and phosphorylation occur on Ser199 of β-actin. However, their roles are not known. To elucidate their roles in diabetic nephropathy, we examined the rat kidney for changes in O-GlcNAcylation of Ser199 (gS199)-actin and in the phosphorylation of Ser199 (pS199)-actin. Both gS199- and pS199-actin molecules had an apparent molecular weight of 40 kDa and were localized as nonfilamentous actin in both the cytoplasm and nucleus. Compared with the normal kidney, the immunostaining intensity of gS199-actin increased in podocytes of the glomeruli and in proximal tubules of the diabetic kidney, whereas that of pS199-actin did not change in podocytes but decreased in proximal tubules. We confirmed that the same results could be observed in the glomeruli of the human diabetic kidney. In podocytes of glomeruli cultured in the presence of the O-GlcNAcase inhibitor Thiamet G, increased O-GlcNAcylation was accompanied by a concomitant decrease in the amount of filamentous actin and in morphological changes. Our present results demonstrate that dysregulation of O-GlcNAcylation and phosphorylation of Ser199 occurred in diabetes, which may contribute partially to the causes of the morphological changes in the glomeruli and tubules. gS199- and pS199-actin will thus be useful for the pathological evaluation of diabetic nephropathy.
Keywords: Goto-Kakizaki rat; O-GlcNAcylation; diabetic nephropathy; phosphorylation and dephosphorylation; β-actin.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures



References
-
- Akimoto Y, Hart GW, Wells L, Vosseller K, Yamamoto K, Munetomo E, Ohara-Imaizumi M, Nishiwaki C, Nagamatsu S, Hirano H, Kawakami H. Elevation of the post-translational modification of proteins by O-linked N-acetylglucosamine leads to deterioration of the glucose-stimulated insulin secretion in the pancreas of diabetic Goto-Kakizaki rats. Glycobiology 17: 127–140, 2007. doi: 10.1093/glycob/cwl067. - DOI - PubMed
-
- Akimoto Y, Miura Y, Toda T, Wolfert MA, Wells L, Boons G-J, Hart GW, Endo T, Kawakami H. Morphological changes in diabetic kidney are associated with increased O-GlcNAcylation of cytoskeletal proteins including α-actinin 4. Clin Proteomics 8: 15, 2011. doi: 10.1186/1559-0275-8-15. - DOI - PMC - PubMed
-
- Akimoto Y, Yamamoto K, Munetomo E, Wells L, Vosseller K, Hart GW, Kawakami H, Hirano H. Elevated post-translational modification of proteins by O-linked N-acetylglucosamine in various tissues of diabetic Goto-Kakizaki rats accompanied by diabetic complications. Acta Histochem Cytochem 38: 131–142, 2005. doi: 10.1267/ahc.38.131. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases