

Causes and consequences of representational drift
- PMID: 31569062
- PMCID: PMC7385530
- DOI: 10.1016/j.conb.2019.08.005
Causes and consequences of representational drift
Abstract
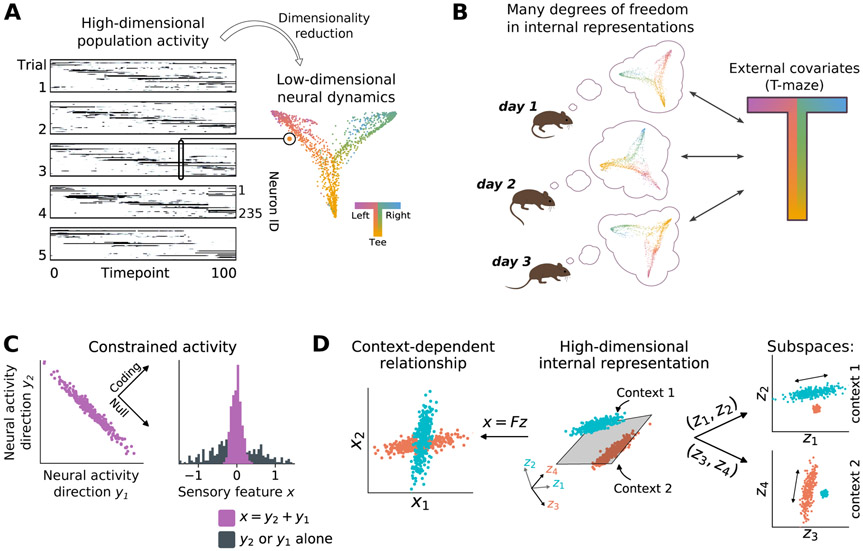
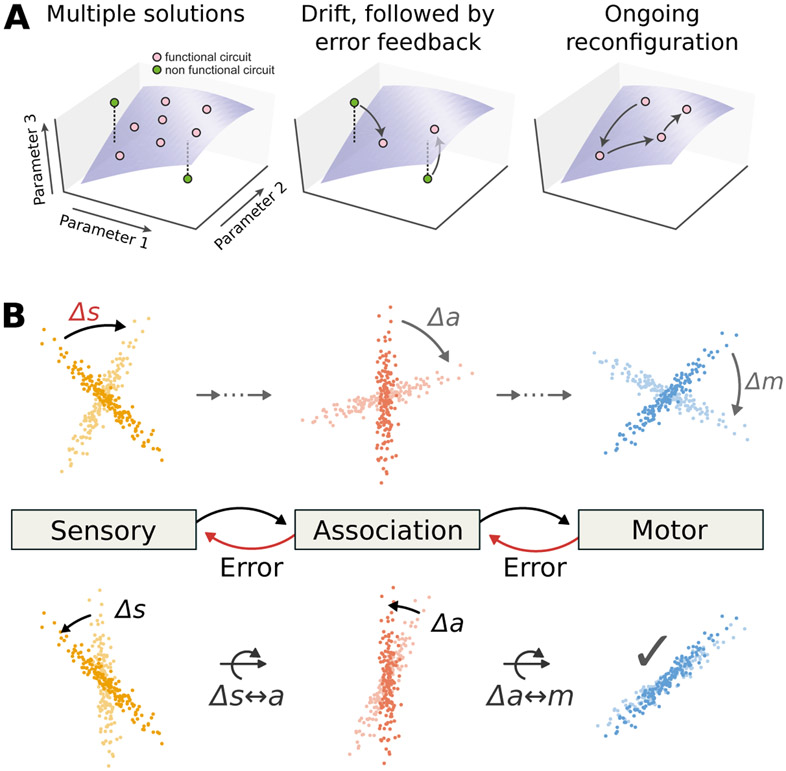
The nervous system learns new associations while maintaining memories over long periods, exhibiting a balance between flexibility and stability. Recent experiments reveal that neuronal representations of learned sensorimotor tasks continually change over days and weeks, even after animals have achieved expert behavioral performance. How is learned information stored to allow consistent behavior despite ongoing changes in neuronal activity? What functions could ongoing reconfiguration serve? We highlight recent experimental evidence for such representational drift in sensorimotor systems, and discuss how this fits into a framework of distributed population codes. We identify recent theoretical work that suggests computational roles for drift and argue that the recurrent and distributed nature of sensorimotor representations permits drift while limiting disruptive effects. We propose that representational drift may create error signals between interconnected brain regions that can be used to keep neural codes consistent in the presence of continual change. These concepts suggest experimental and theoretical approaches to studying both learning and maintenance of distributed and adaptive population codes.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Figures
References
-
- Tonegawa Susumu, Pignatelli Michele, Roy Dheeraj S, and Ryan Tomas J. Memory engram storage and retrieval. Current opinion in neurobiology, 35:101–109, 2015. - PubMed
-
- Mongillo Gianluigi, Rumpel Simon, and Loewenstein Yonatan. Intrinsic volatility of synaptic connections—a challenge to the synaptic trace theory of memory. Current opinion in neurobiology, 46:7–13, 2017. - PubMed
-
- Rumpel Simon and Triesch Jochen. The dynamic connectome. e-Neuroforum, 22(3):48–53, 2016.
-
-
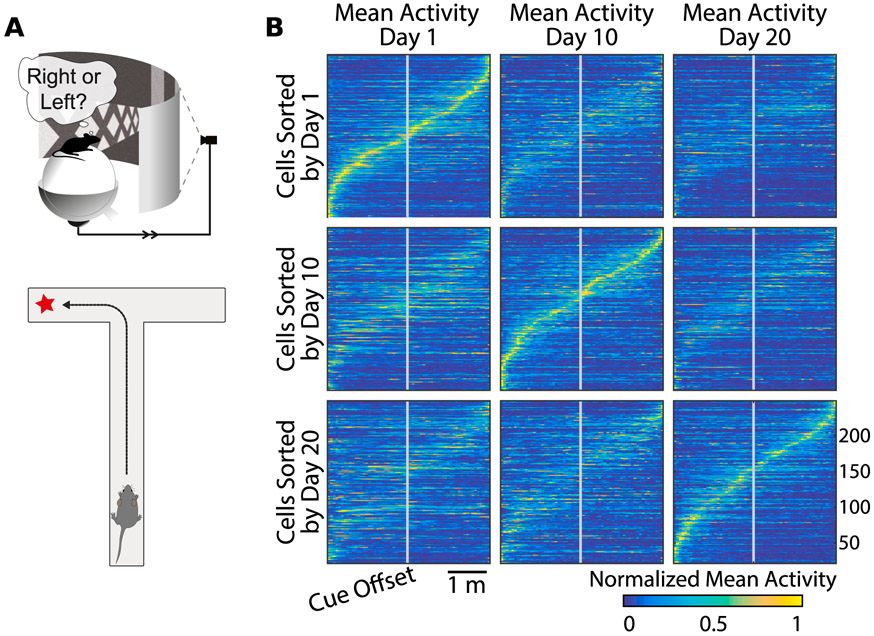
Driscoll Laura N, Pettit Noah L, Minderer Matthias, Chettih Selmaan N, and Harvey Christopher D. Dynamic reorganization of neuronal activity patterns in parietal cortex. Cell, 170(5):986–999, 2017.
The authors examine neural representations for spatial navigation in mouse posterior parietal cortex using a closed-loop virtual reality environment, and find that the neural code drifts and reconfigures itself over days.
-
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
