

Store-Operated Calcium Channels: From Function to Structure and Back Again
- PMID: 31570335
- PMCID: PMC7103539
- DOI: 10.1101/cshperspect.a035055
Store-Operated Calcium Channels: From Function to Structure and Back Again
Abstract
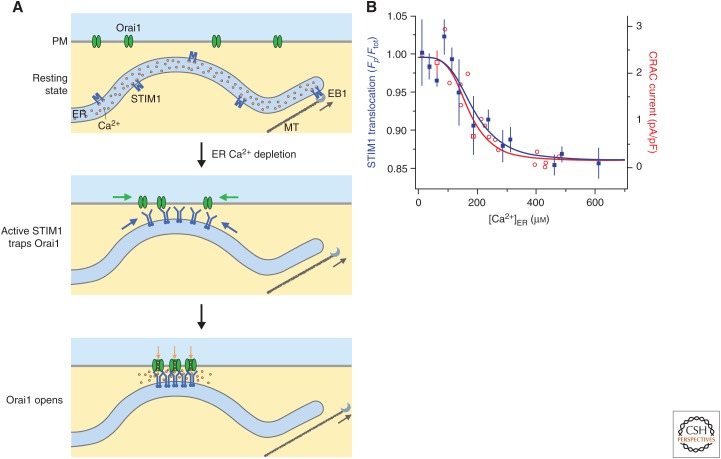
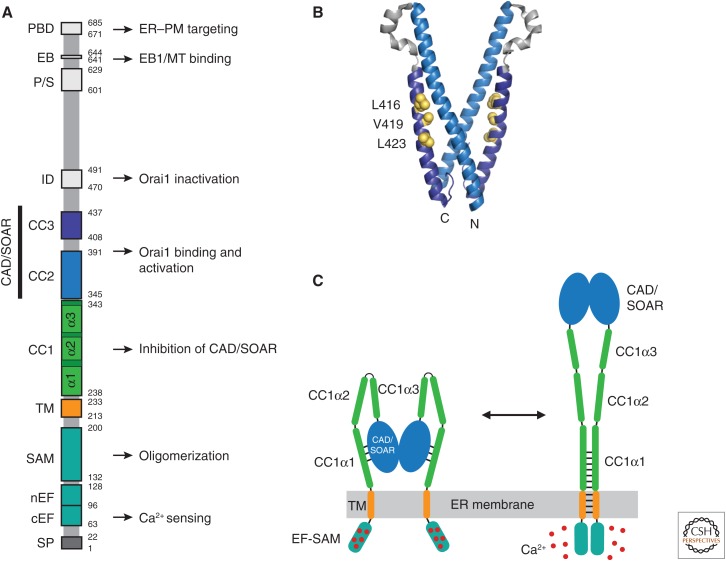
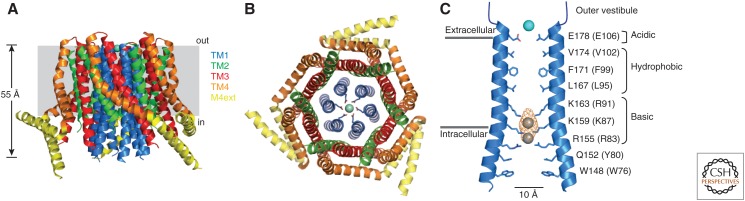
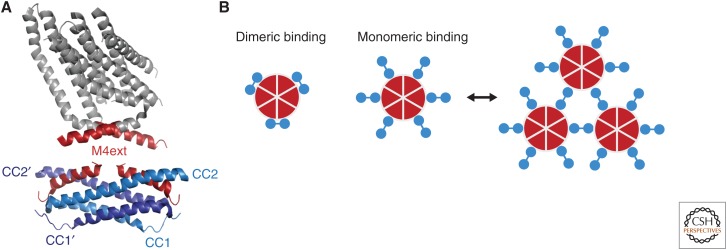
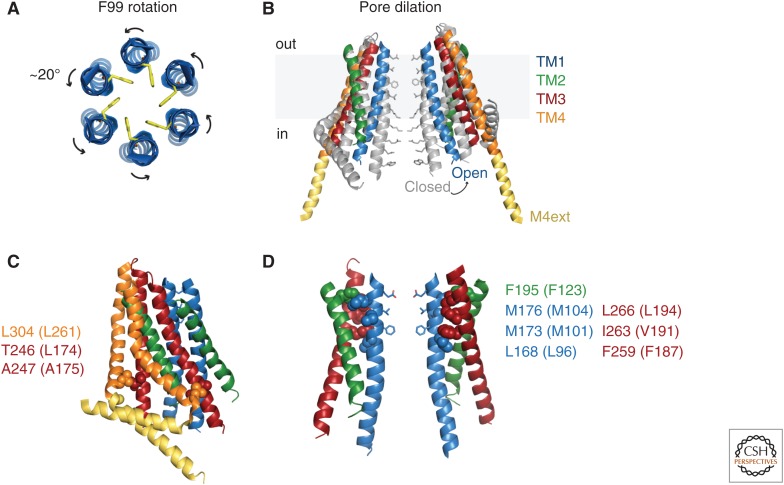
Store-operated calcium (Ca2+) entry (SOCE) occurs through a widely distributed family of ion channels activated by the loss of Ca2+ from the endoplasmic reticulum (ER). The best understood of these is the Ca2+ release-activated Ca2+ (CRAC) channel, which is notable for its unique activation mechanism as well as its many essential physiological functions and the diverse pathologies that result from dysregulation. In response to ER Ca2+ depletion, CRAC channels are formed through a diffusion trap mechanism at ER-plasma membrane (PM) junctions, where the ER Ca2+-sensing stromal interaction molecule (STIM) proteins bind and activate hexamers of Orai pore-forming proteins to trigger Ca2+ entry. Cell biological studies are clarifying the architecture of ER-PM junctions, their roles in Ca2+ and lipid transport, and functional interactions with cytoskeletal proteins. Molecular structures of STIM and Orai have inspired a multitude of mutagenesis and electrophysiological studies that reveal potential mechanisms for how STIM is toggled between inactive and active states, how it binds and activates Orai, and the importance of STIM-binding stoichiometry for opening the channel and establishing its signature characteristics of extremely high Ca2+ selectivity and low Ca2+ conductance.
Copyright © 2020 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous