

Hepatitis C Virus Replication
- PMID: 31570388
- PMCID: PMC7050578
- DOI: 10.1101/cshperspect.a037093
Hepatitis C Virus Replication
Abstract
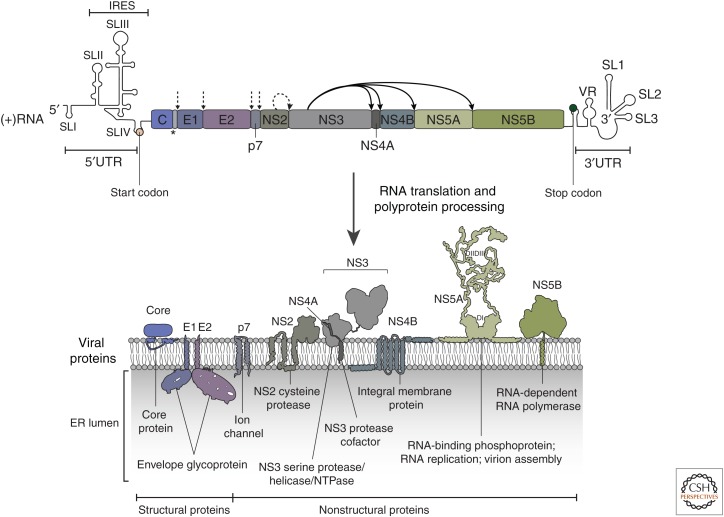
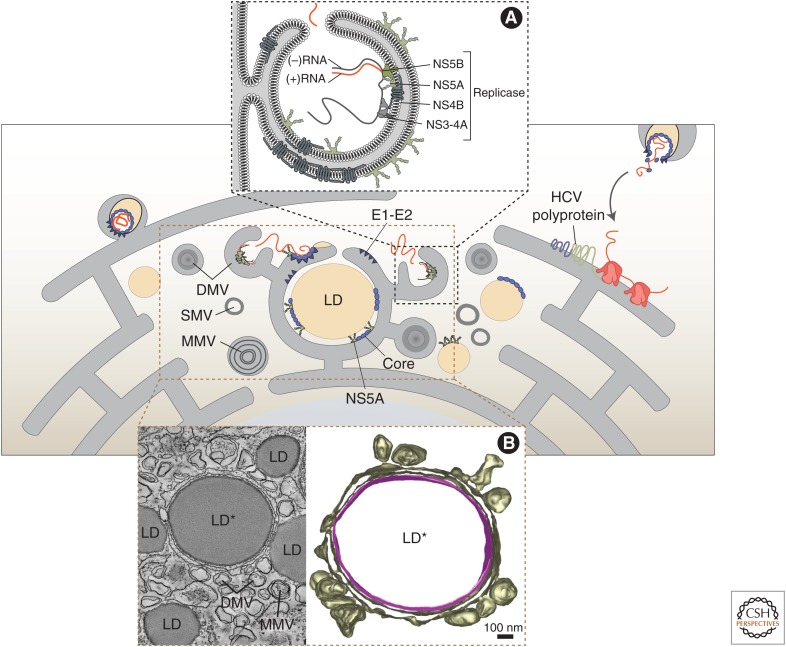
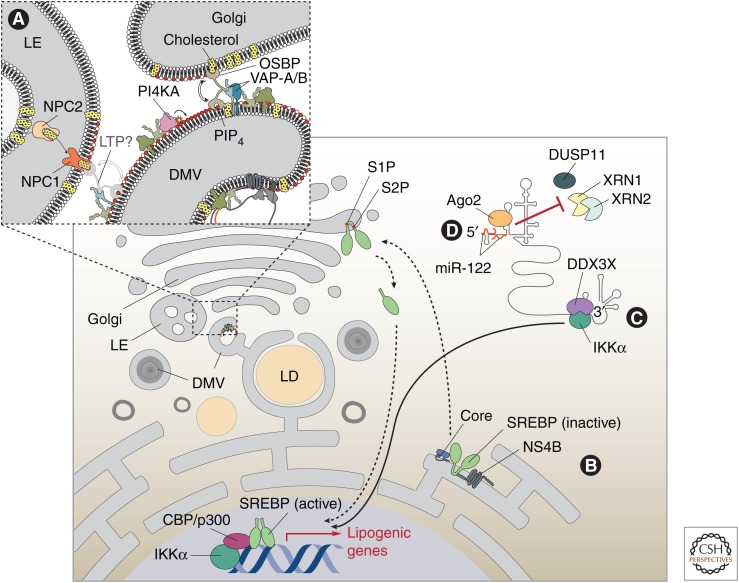
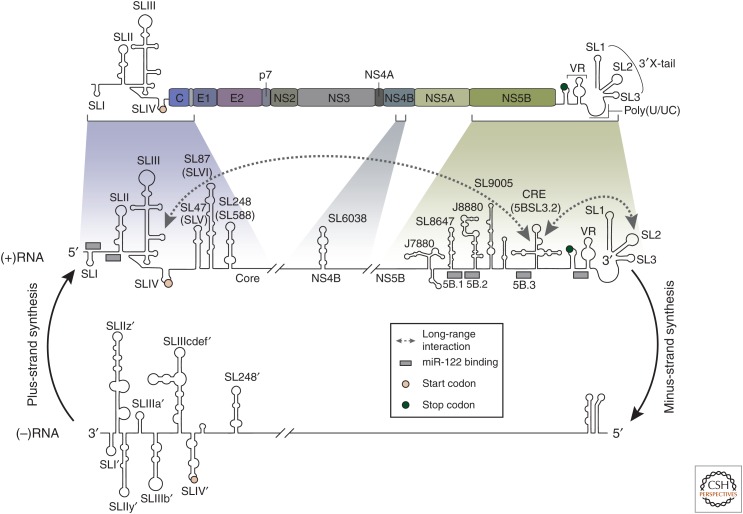
Replication and amplification of the viral genome is a key process for all viruses. For hepatitis C virus (HCV), a positive-strand RNA virus, amplification of the viral genome requires the synthesis of a negative-sense RNA template, which is in turn used for the production of new genomic RNA. This process is governed by numerous proteins, both host and viral, as well as distinct lipids and specific RNA elements within the positive- and negative-strand RNAs. Moreover, this process requires specific changes to host cell ultrastructure to create microenvironments conducive to viral replication. This review will focus on describing the processes and factors involved in facilitating or regulating HCV genome replication.
Copyright © 2020 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
-
- Amador-Cañizares Y, Panigrahi M, Huys A, Kunden RD, Adams HM, Schinold MJ, Wilson JA. 2018b. miR-122, small RNA annealing and sequence mutations alter the predicted structure of the hepatitis C virus 5′ UTR RNA to stabilize and promote viral RNA accumulation. Nucleic Acids Res 46: 9776–9792. 10.1093/nar/gky662 - DOI - PMC - PubMed
-
- Appel N, Pietschmann T, Bartenschlager R. 2005. Mutational analysis of hepatitis C virus nonstructural protein 5A: potential role of differential phosphorylation in RNA replication and identification of a genetically flexible domain. J Virol 79: 3187–3194. 10.1128/JVI.79.5.3187-3194.2005 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources