

Dissociation of mitochondrial HK-II elicits mitophagy and confers cardioprotection against ischemia
- PMID: 31570704
- PMCID: PMC6768853
- DOI: 10.1038/s41419-019-1965-7
Dissociation of mitochondrial HK-II elicits mitophagy and confers cardioprotection against ischemia
Abstract
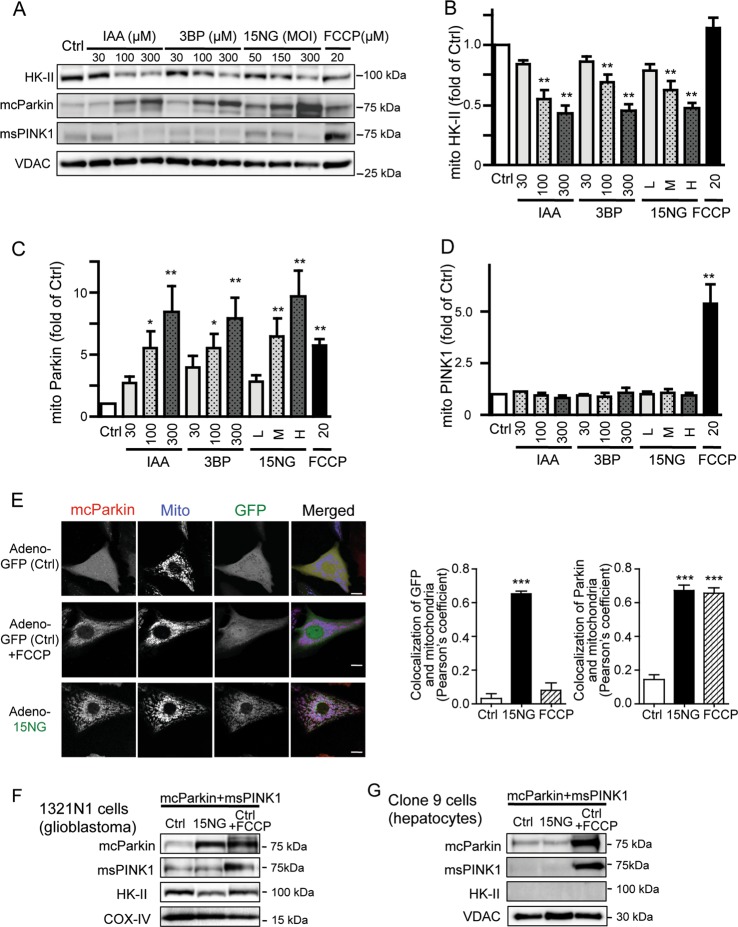
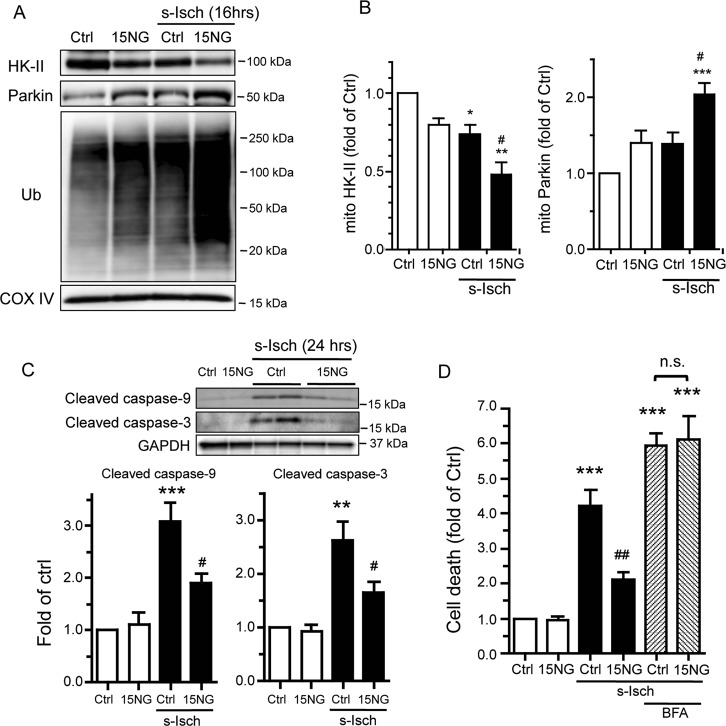
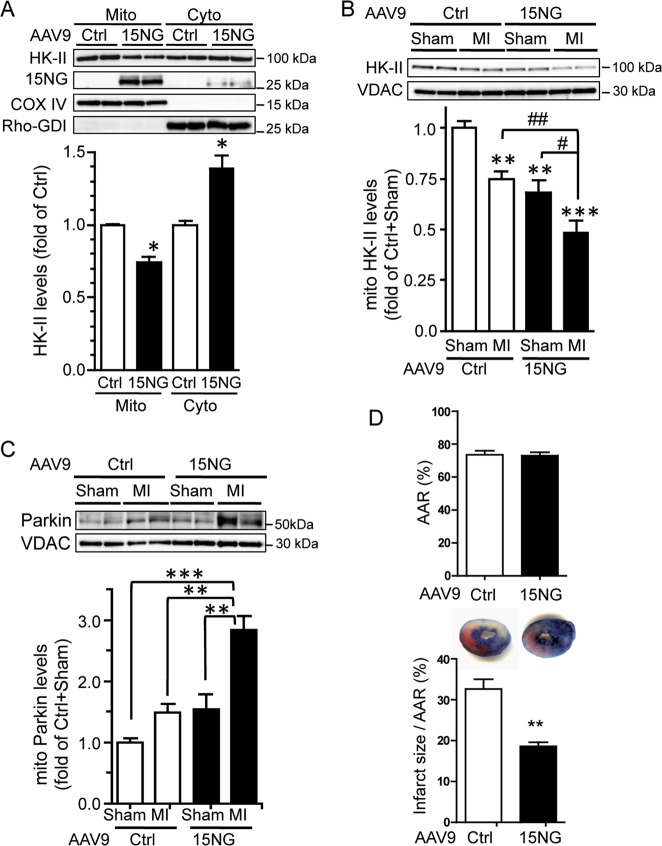
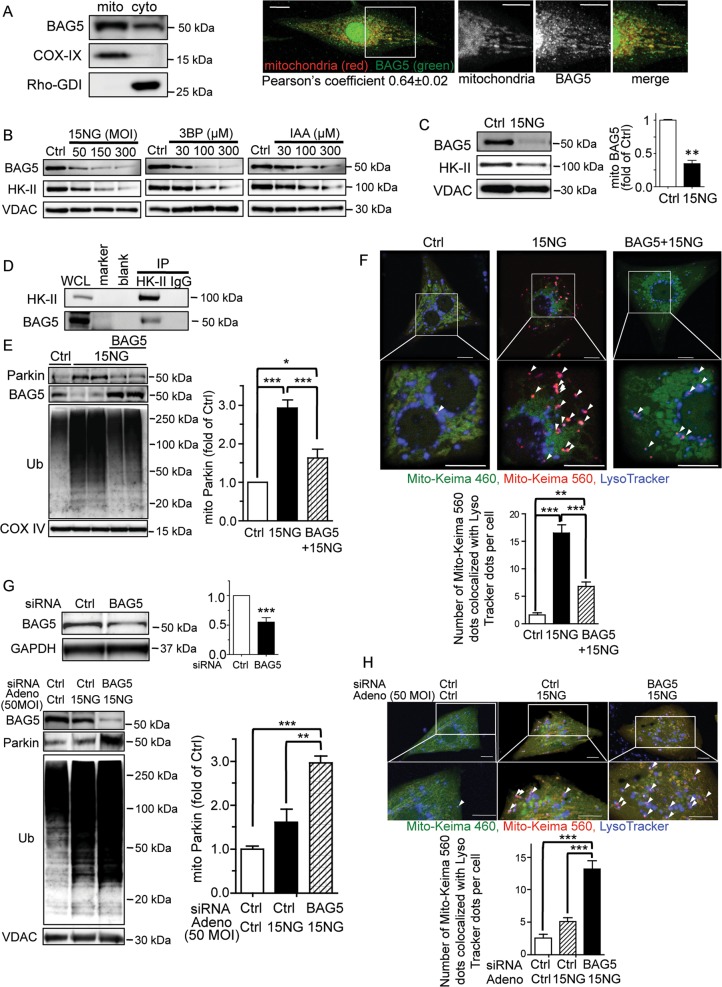
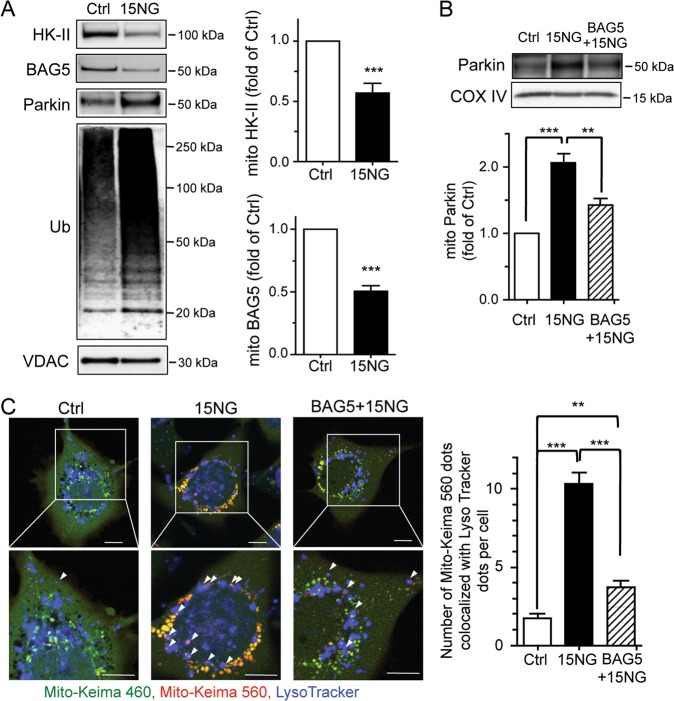
Preservation of mitochondrial integrity is critical for maintaining cellular homeostasis. Mitophagy is a mitochondria-specific type of autophagy which eliminates damaged mitochondria thereby contributing to mitochondrial quality control. Depolarization of the mitochondrial membrane potential is an established mechanism for inducing mitophagy, mediated through PINK1 stabilization and Parkin recruitment to mitochondria. Hexokinase-II (HK-II) which catalyzes the first step in glucose metabolism, also functions as a signaling molecule to regulate cell survival, and a significant fraction of cellular HK-II is associated with mitochondria (mitoHK-II). We demonstrate here that pharmacological interventions and adenoviral expression of a mitoHK-II dissociating peptide which reduce mitoHK-II levels lead to robust increases in mitochondrial Parkin and ubiquitination of mitochondrial proteins in cardiomyocytes and in a human glioblastoma cell line 1321N1, independent of mitochondrial membrane depolarization or PINK1 accumulation. MitoHK-II dissociation-induced mitophagy was demonstrated using Mito-Keima in cardiomyocytes and in 1321N1 cells. Subjecting cardiomyocytes or the in vivo heart to ischemia leads to modest dissociation of mitoHK-II. This response is potentiated by expression of the mitoHK-II dissociating peptide, which increases Parkin recruitment to mitochondria and, importantly, provides cardioprotection against ischemic stress. These results suggest that mitoHK-II dissociation is a physiologically relevant cellular event that is induced by ischemic stress, the enhancement of which protects against ischemic damage. The mechanism which underlies the effects of mitoHK-II dissociation can be attributed to the ability of Bcl2-associated athanogene 5 (BAG5), an inhibitor of Parkin, to localize to mitochondria and form a molecular complex with HK-II. Overexpression of BAG5 attenuates while knockdown of BAG5 sensitizes the effect of mitoHK-II dissociation on mitophagy. We suggest that HK-II, a glycolytic molecule, can function as a sensor for metabolic derangements at mitochondria to trigger mitophagy, and modulating the intracellular localization of HK-II could be a novel way of regulating mitophagy to prevent cell death induced by ischemic stress.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
