

doi: 10.1038/s41588-019-0498-4.
Epub 2019 Sep 30.
Postzygotic inactivating mutations of RHOA cause a mosaic neuroectodermal syndrome
Affiliations
- PMID: 31570889
- PMCID: PMC6858542
- DOI: 10.1038/s41588-019-0498-4
Item in Clipboard
Postzygotic inactivating mutations of RHOA cause a mosaic neuroectodermal syndrome
Nat Genet.
2019 Oct.
Erratum in
-
Author Correction: Postzygotic inactivating mutations of RHOA cause a mosaic neuroectodermal syndrome.Nat Genet. 2019 Nov;51(11):1660. doi: 10.1038/s41588-019-0527-3. Nat Genet. 2019. PMID: 31611689
-
Author Correction: Postzygotic inactivating mutations of RHOA cause a mosaic neuroectodermal syndrome.Nat Genet. 2020 Mar;52(3):353. doi: 10.1038/s41588-019-0565-x. Nat Genet. 2020. PMID: 32034319
Abstract
Hypopigmentation along Blaschko's lines is a hallmark of a poorly defined group of mosaic syndromes whose genetic causes are unknown. Here we show that postzygotic inactivating mutations of RHOA cause a neuroectodermal syndrome combining linear hypopigmentation, alopecia, apparently asymptomatic leukoencephalopathy, and facial, ocular, dental and acral anomalies. Our findings pave the way toward elucidating the etiology of pigmentary mosaicism and highlight the role of RHOA in human development and disease.
Conflict of interest statement
COMPETING FINANCIAL INTERESTS
The authors declare no competing financial interests.
Figures
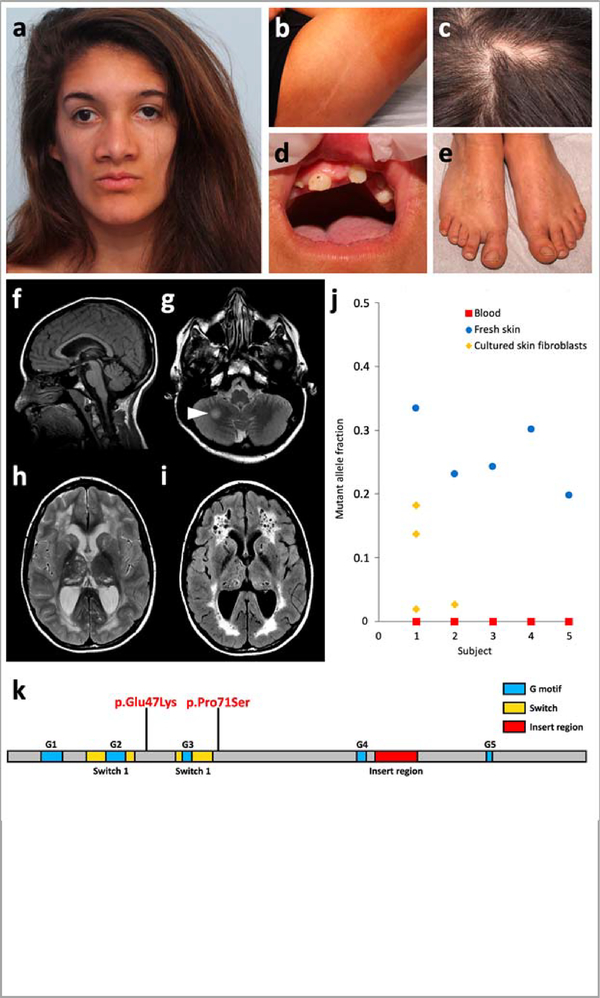
a-e, Craniofacial appearance, linear hypopigmentation and other extracutaneous anomalies in subject S1. f-i, Brain MRI of subject S1 at 15 years. A second MRI, conducted 6 months later, did not show any significant change. Subjects S2 and S4 had similar but milder MRI abnormalities, including enlarged temporal horns of the lateral ventricles (Supplementary Fig. 2). Sagittal T1-weighted image revealed preserved midline structures (f). Axial T2-weighted images revealed a focal hyperintense lesion in the right hemisphere of the cerebellum (g, arrowhead), and diffuse cystic leukoencephalopathy with mildly enlarged lateral ventricles and cysts in the thalami and caudate nuclei (h). The leukoencephalopathy and presence of multiple cysts is confirmed on fluid-attenuated inversion recovery (FLAIR) sequences (i). j, Mutant allele fraction of RHOA mutations in the five subjects studied in WES or TUDS. k, Linear representation of RHOA and localization of the two mutations.
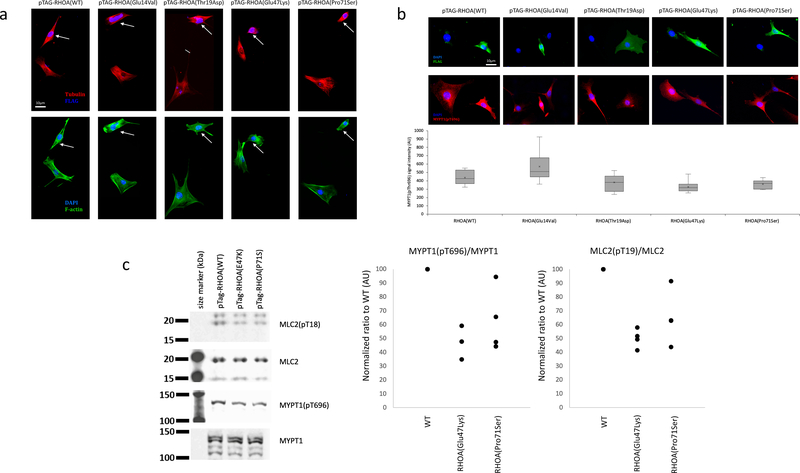
a,b, Cytoskeletal organization and morphology in NIH/3T3 cells transfected with wild-type, constitutively active (p.Gly14Val), dominant-negative (p.Thr19Asn), (p.Glu47Lys) and (p.Pro71Ser) forms of RHOA. a, Up: Cells transfected with wild-type RHOA or p.Gly14Val mutant display expected increase in F-actin staining, particularly with regard to stress fibers which are brighter, thicker and more numerous. Cells transfected with p.Thr19Asn, p.Glu47Lys or (p.Pro71Ser) mutants barely contain any stress fibers at all. Low: Dual labeling for DAPI (blue) and alpha-tubulin (green) does not reveal significant differences in the gross organization of microtubule cytoskeleton or nuclear morphology between different mutants. All cells (n = 20 per group) selected at random across the cover slip that were individually examined showed reduced stress fibers and limited cell spreading. b, Up: FLAG staining does not reveal any visible differences in the subcellular localization of different RhoA mutants. All mutants tend to impair cell spreading, while wild-type RhoA overexpressing cells maintain normal morphology. Middle: Dual labeling for DAPI (blue) and pMYPT1 (red) shows decrease in signal intensity of MYPT1(pT696) staining upon transfection with T19N (control), E47K or P71S mutants. Low: Quantification of MYPT1(pThr696) staining shows significant decrease (n = 20, 24, 14, 30, and 13 cells, for WT, G14V, T19N, E47K, and P71S, respectively). Box plot elements: 5°, 25°, median, mean (cross), 75° and 95°percentiles. c, Levels of phosphorylated MYPT1(pThr696) and MLC2(pThr19). Left: Cropped images of Western blot experiment showing expression levels of total MYPT1, phospho-MYPT1, total MLC2, and phospho-MLC2. There is a visible reduction in phospho-MYPT1 and phospho-MLC2 when RhoA(Thr19Asn) or RhoA(Glu47Lys) are overexpressed. Middle and right: dot plot of normalized ratio (4 independent experiments) for phospho-MYPT1 and phospho-MLC2 normalized to total MYPT1 and MLC2, respectively, indicate reduction in MYPT1(pThr696) and MLC2(pThr19) upon RhoA(Glu47Lys) or RhoA(Pro71Ser) overexpression. Further analyses for p.(Glu47Lys) are shown in Supplementary Figure 7. Full scans of blots are provided in Supplementary Figure 8.
References
-
- Sybert VP Hypomelanosis of Ito: a description, not a diagnosis. J. Invest. Dermatol 103, S141–S143 (1994). - PubMed
-
- Pavone P, Praticò AD, Ruggieri M & Falsaperla R Hypomelanosis of Ito: a round on the frequency and type of epileptic complications. Neurol. Sci 36, 1173–1180 (2015). - PubMed
-
- Hall A Rho GTPases and the actin cytoskeleton. Science 279, 509–514 (1998). - PubMed
-
- Vetter IR & Wittinghofer A The guanine nucleotide-binding switch in three dimensions. Science 294, 1299–1304 (2001). - PubMed
Methods-only References
-
- Thevenon J et al. Diagnostic odyssey in severe neurodevelopmental disorders: toward clinical whole-exome sequencing as a first-line diagnostic test. Clin. Genet 89, 700–707 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
