

Low- and high-thermogenic brown adipocyte subpopulations coexist in murine adipose tissue
- PMID: 31573981
- PMCID: PMC6934193
- DOI: 10.1172/JCI129167
Low- and high-thermogenic brown adipocyte subpopulations coexist in murine adipose tissue
Abstract
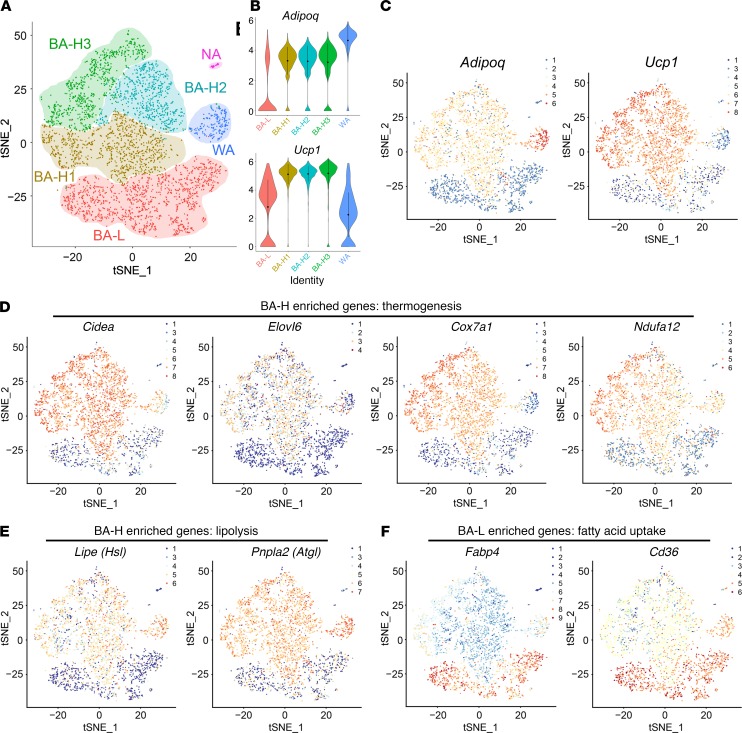
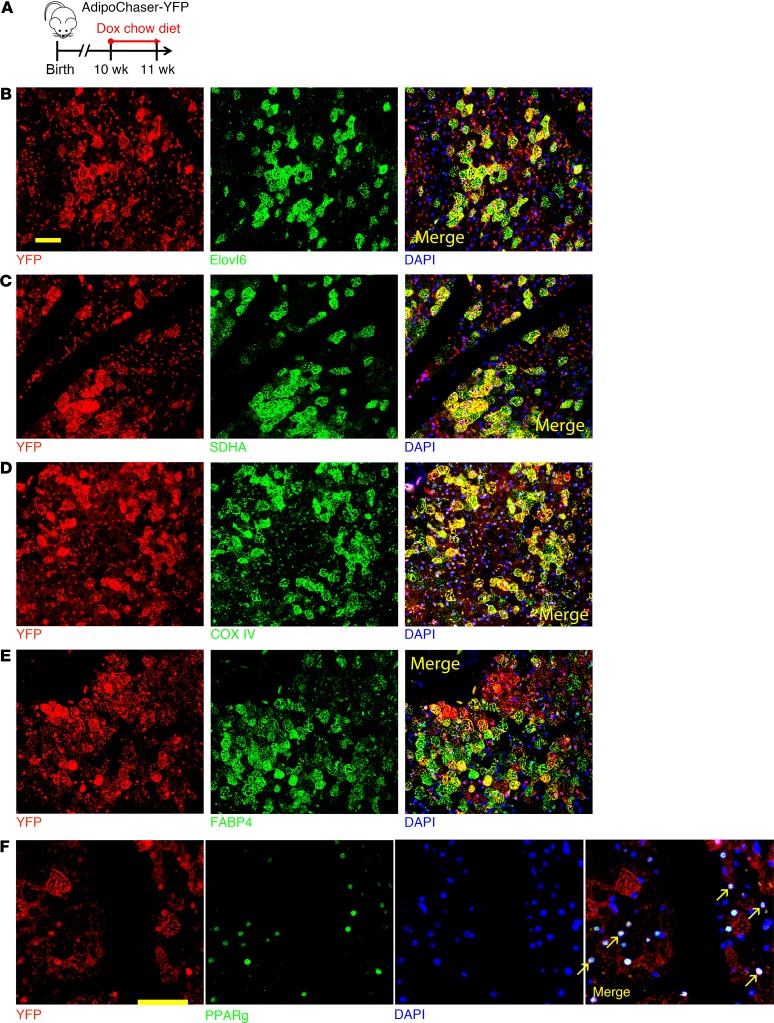
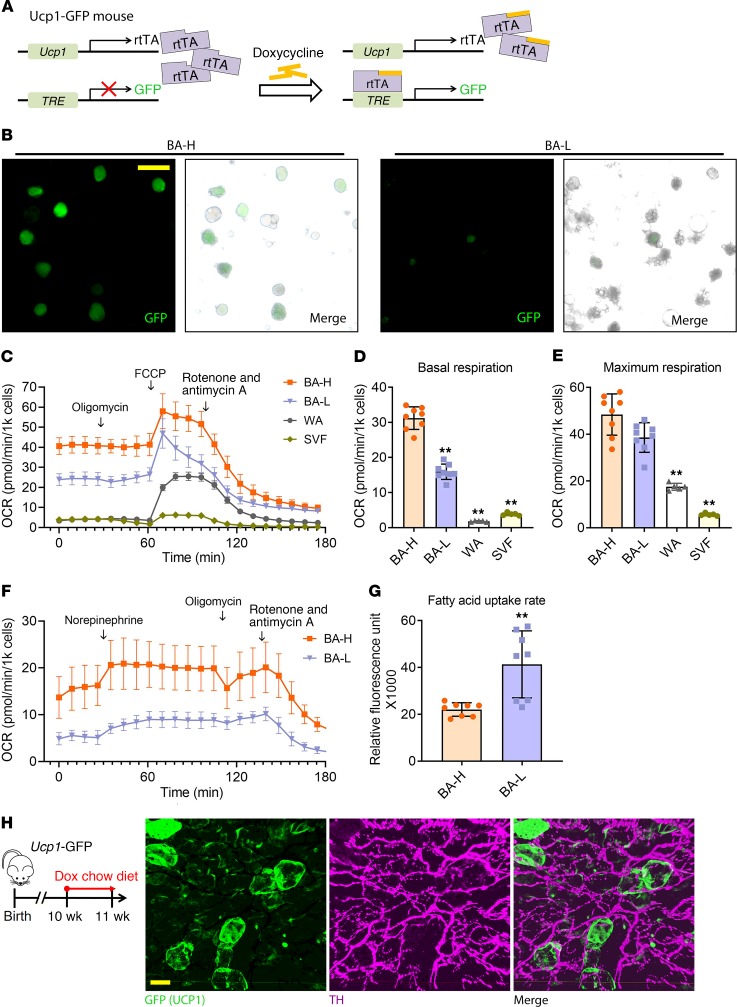
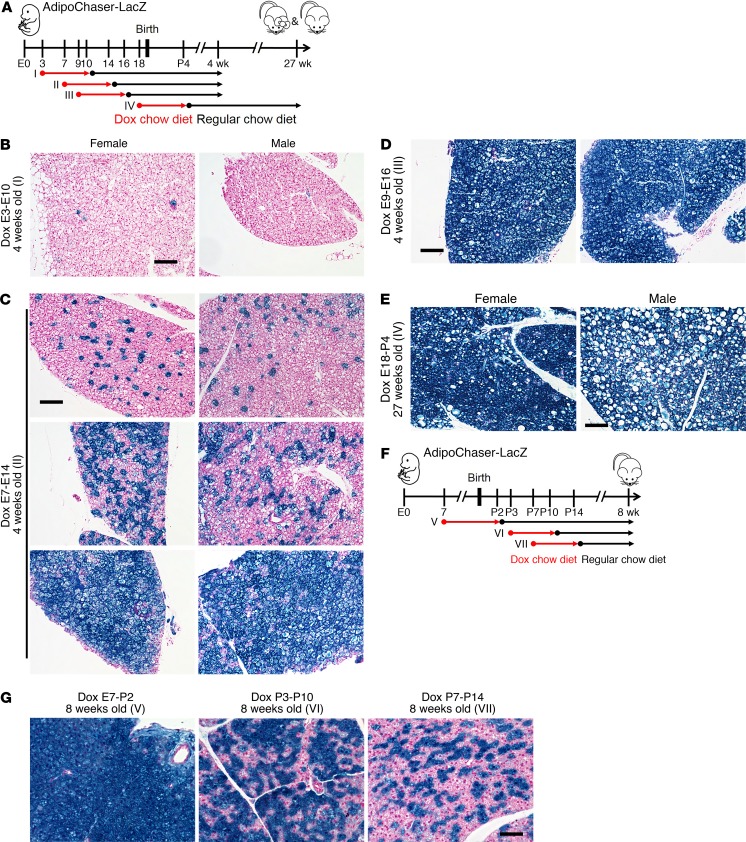
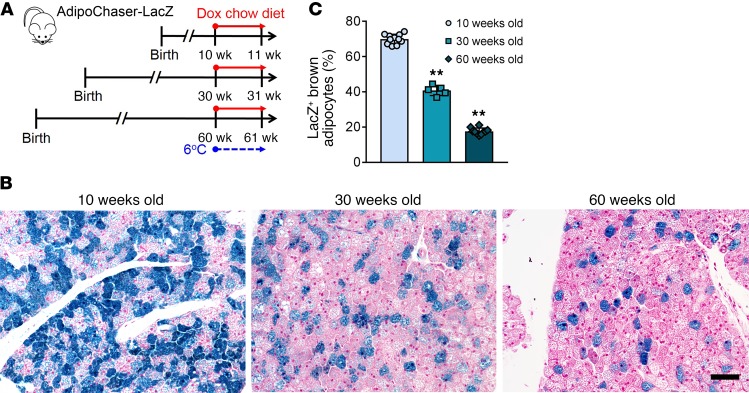
Brown adipose tissue (BAT), as the main site of adaptive thermogenesis, exerts beneficial metabolic effects on obesity and insulin resistance. BAT has been previously assumed to contain a homogeneous population of brown adipocytes. Utilizing multiple mouse models capable of genetically labeling different cellular populations, as well as single-cell RNA sequencing and 3D tissue profiling, we discovered a brown adipocyte subpopulation with low thermogenic activity coexisting with the classical high-thermogenic brown adipocytes within the BAT. Compared with the high-thermogenic brown adipocytes, these low-thermogenic brown adipocytes had substantially lower Ucp1 and Adipoq expression, larger lipid droplets, and lower mitochondrial content. Functional analyses showed that, unlike the high-thermogenic brown adipocytes, the low-thermogenic brown adipocytes have markedly lower basal mitochondrial respiration, and they are specialized in fatty acid uptake. Upon changes in environmental temperature, the 2 brown adipocyte subpopulations underwent dynamic interconversions. Cold exposure converted low-thermogenic brown adipocytes into high-thermogenic cells. A thermoneutral environment had the opposite effect. The recruitment of high-thermogenic brown adipocytes by cold stimulation is not affected by high-fat diet feeding, but it does substantially decline with age. Our results revealed a high degree of functional heterogeneity of brown adipocytes.
Keywords: Adipose tissue; Metabolism.
Conflict of interest statement
Figures


Comment in
-
Cellular heterogeneity in brown adipose tissue.J Clin Invest. 2020 Jan 2;130(1):65-67. doi: 10.1172/JCI133786. J Clin Invest. 2020. PMID: 31763995 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
