

Stimulator of interferon genes (STING) activation exacerbates experimental colitis in mice
- PMID: 31582793
- PMCID: PMC6776661
- DOI: 10.1038/s41598-019-50656-5
Stimulator of interferon genes (STING) activation exacerbates experimental colitis in mice
Abstract
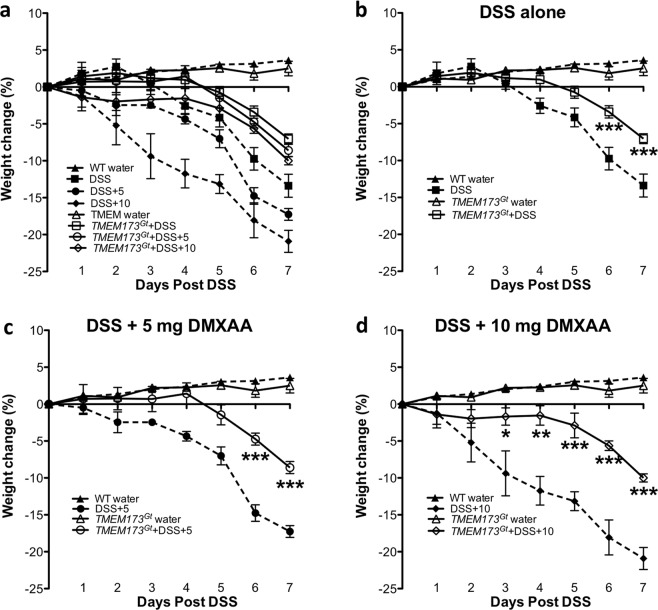
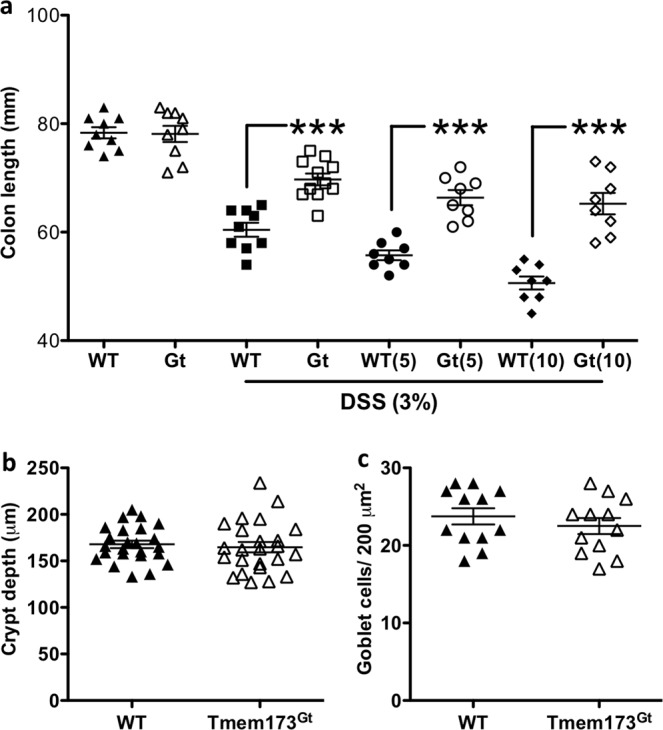
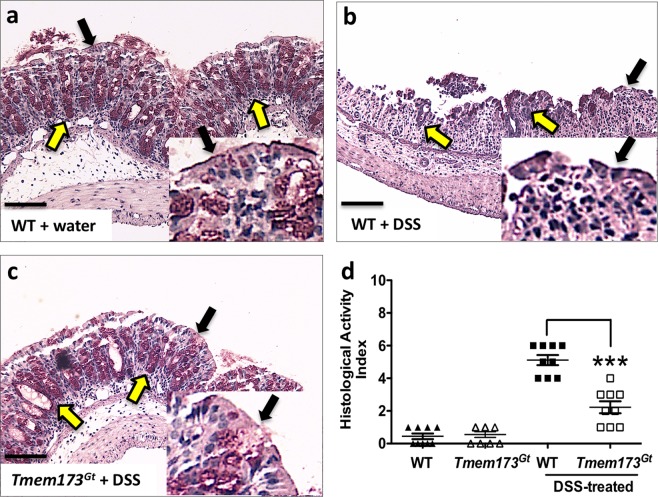
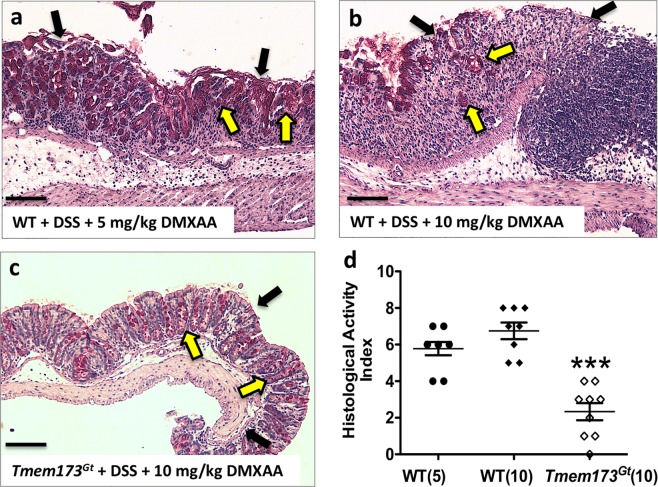
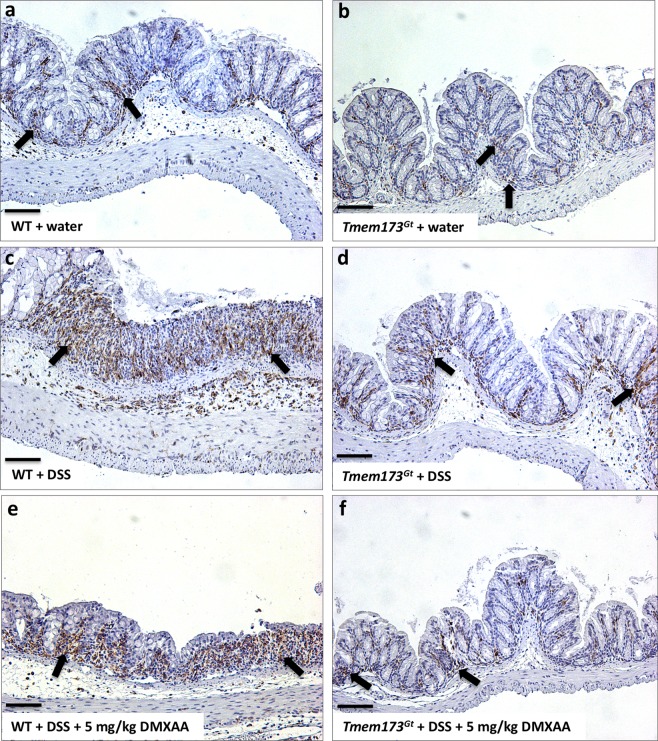
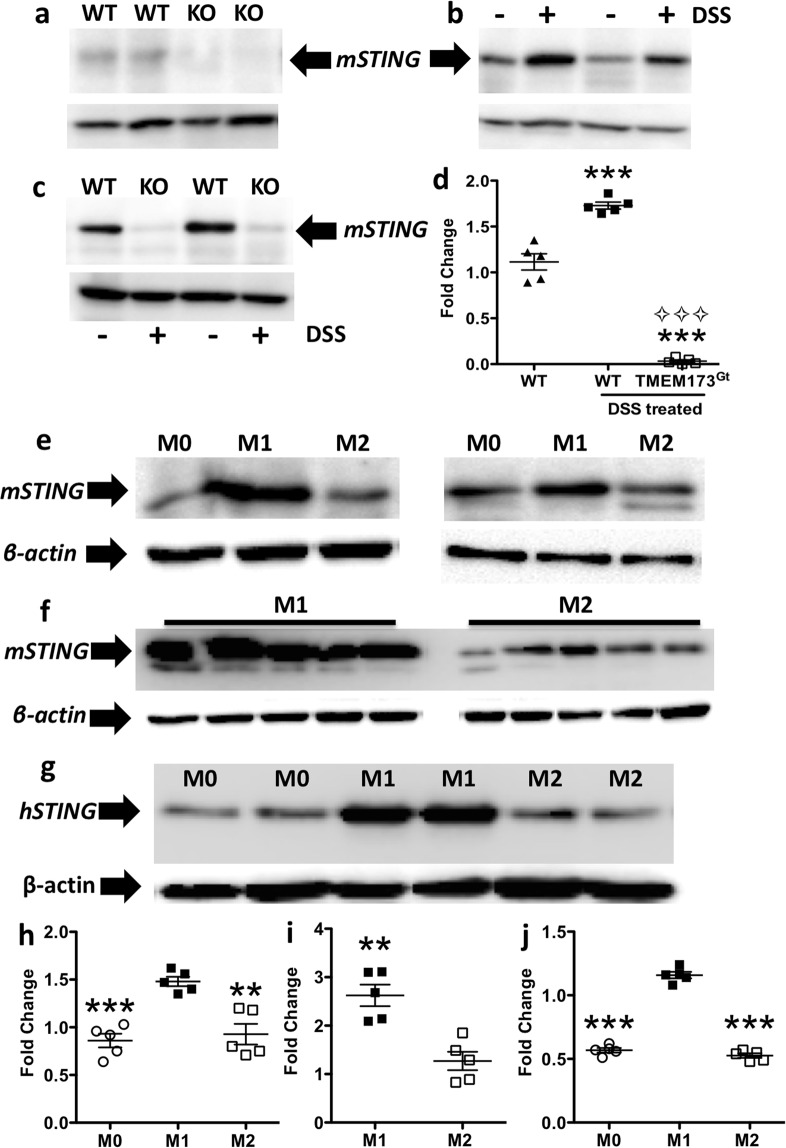
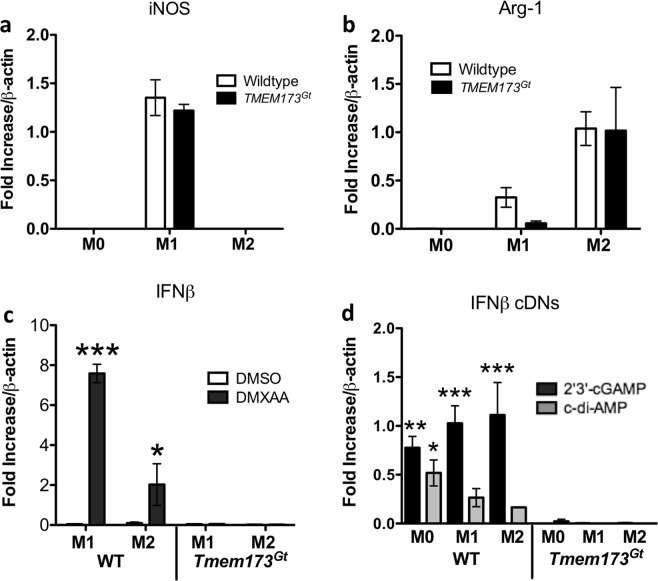
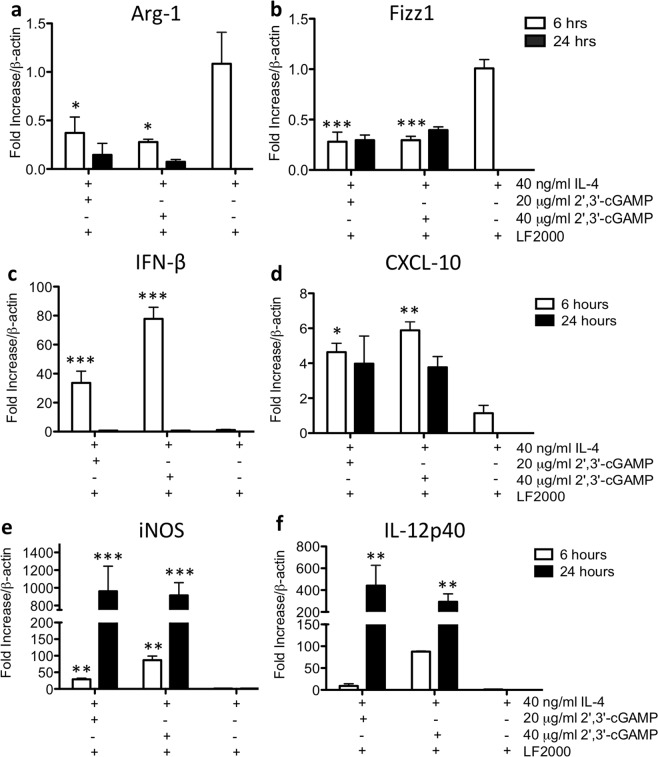
Detection of cytoplasmic DNA by the host's innate immune system is essential for microbial and endogenous pathogen recognition. In mammalian cells, an important sensor is the stimulator of interferon genes (STING) protein, which upon activation by bacterially-derived cyclic dinucleotides (cDNs) or cytosolic dsDNA (dsDNA), triggers type I interferons and pro-inflammatory cytokine production. Given the abundance of bacterially-derived cDNs in the gut, we determined whether STING deletion, or stimulation, acts to modulate the severity of intestinal inflammation in the dextran sodium sulphate (DSS) model of colitis. DSS was administered to Tmem173gt (STING-mutant) mice and to wild-type mice co-treated with DSS and a STING agonist. Colitis severity was markedly reduced in the DSS-treated Tmem173gt mice and greatly exacerbated in wild-type mice co-treated with the STING agonist. STING expression levels were also assessed in colonic tissues, murine bone marrow derived macrophages (BMDMs), and human THP-1 cells. M1 and M2 polarized THP-1 and murine BMDMs were also stimulated with STING agonists and ligands to assess their responses. STING expression was increased in both murine and human M1 polarized macrophages and a STING agonist repolarized M2 macrophages towards an M1-like subtype. Our results suggest that STING is involved in the host's response to acutely-induced colitis.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
