

A mechanism for the activation of the mechanosensitive Piezo1 channel by the small molecule Yoda1
- PMID: 31582801
- PMCID: PMC6776524
- DOI: 10.1038/s41467-019-12501-1
A mechanism for the activation of the mechanosensitive Piezo1 channel by the small molecule Yoda1
Abstract
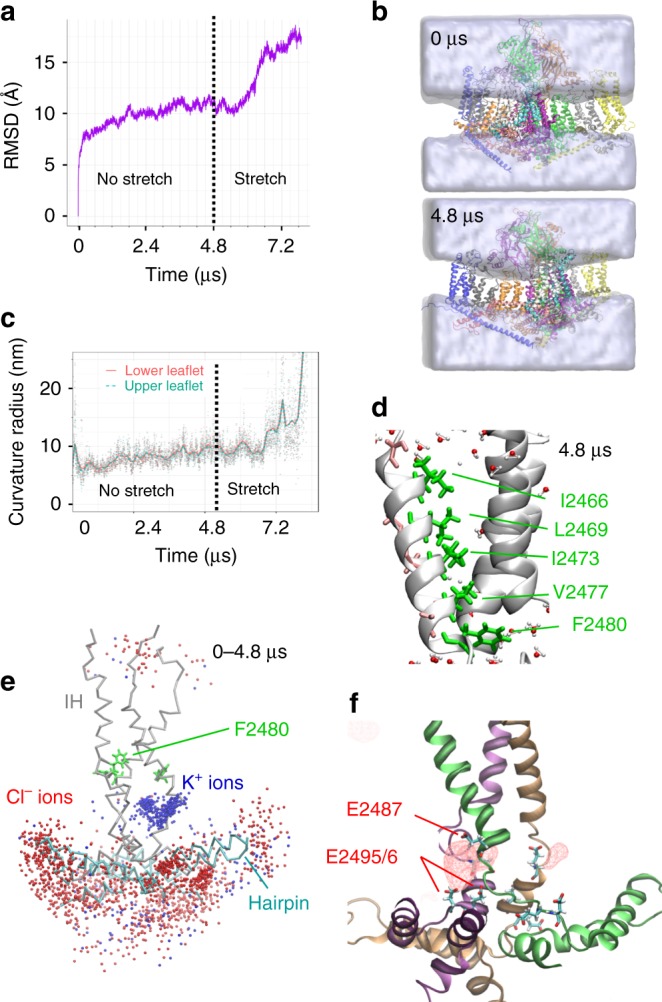
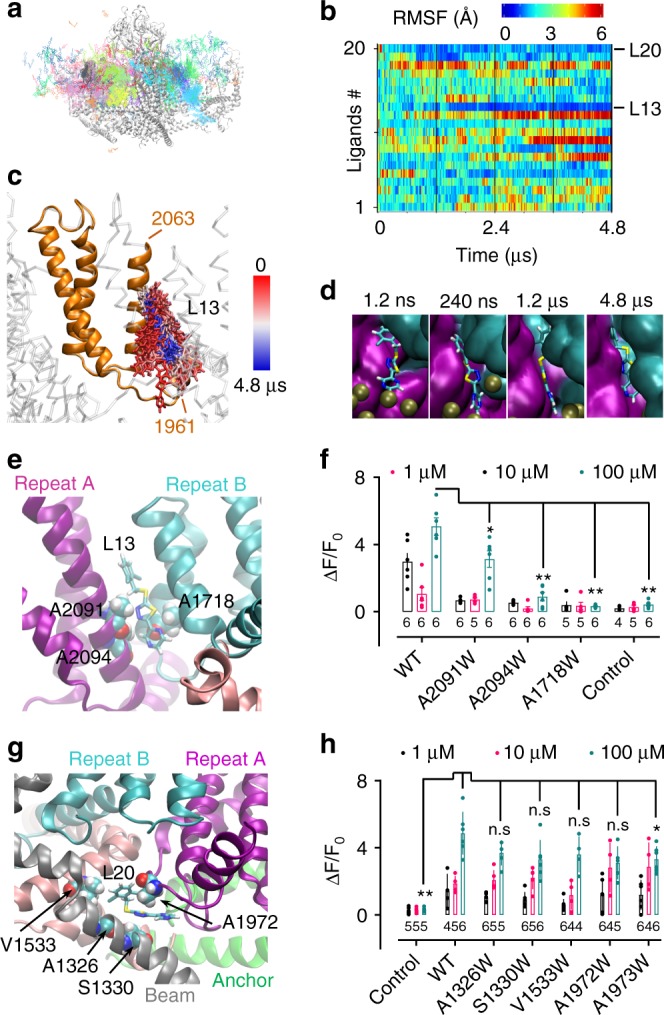
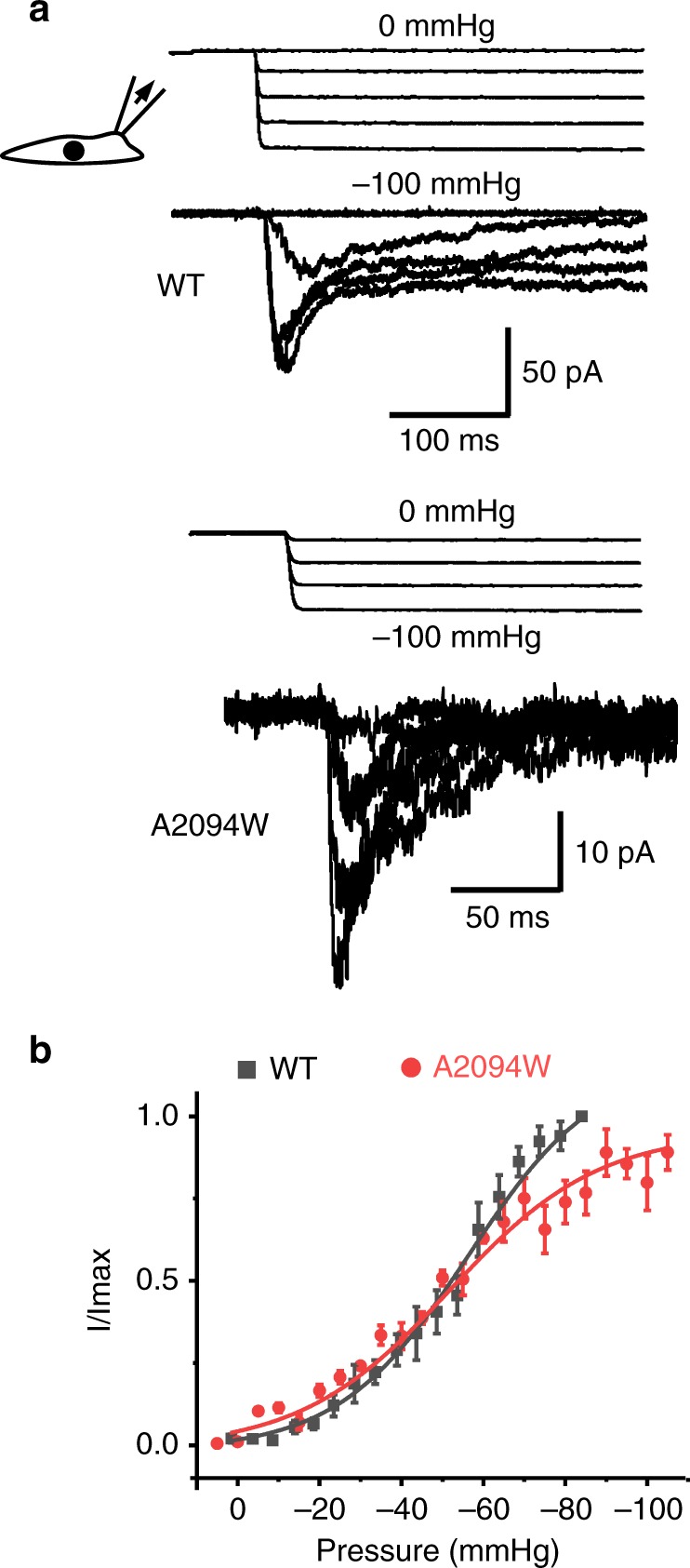
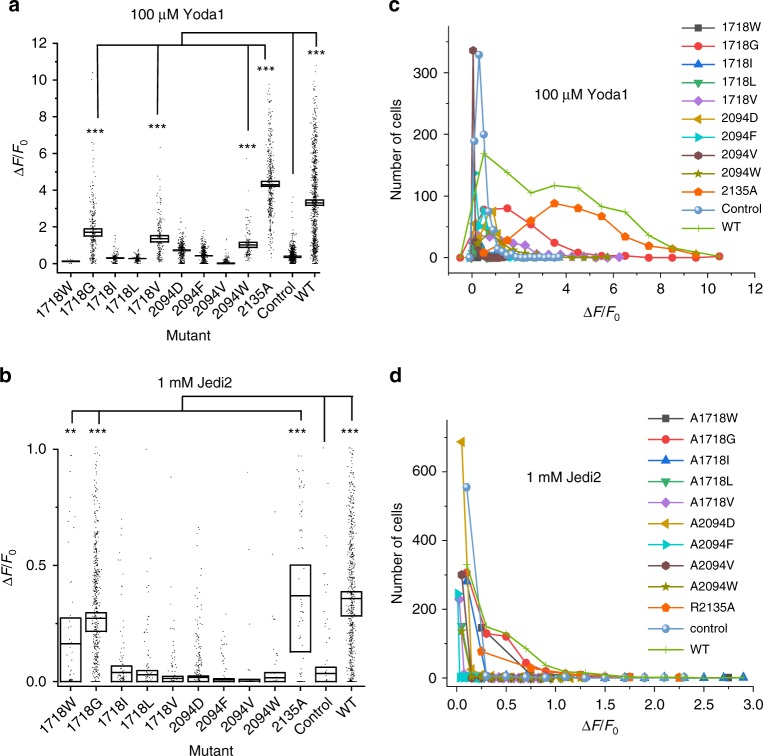
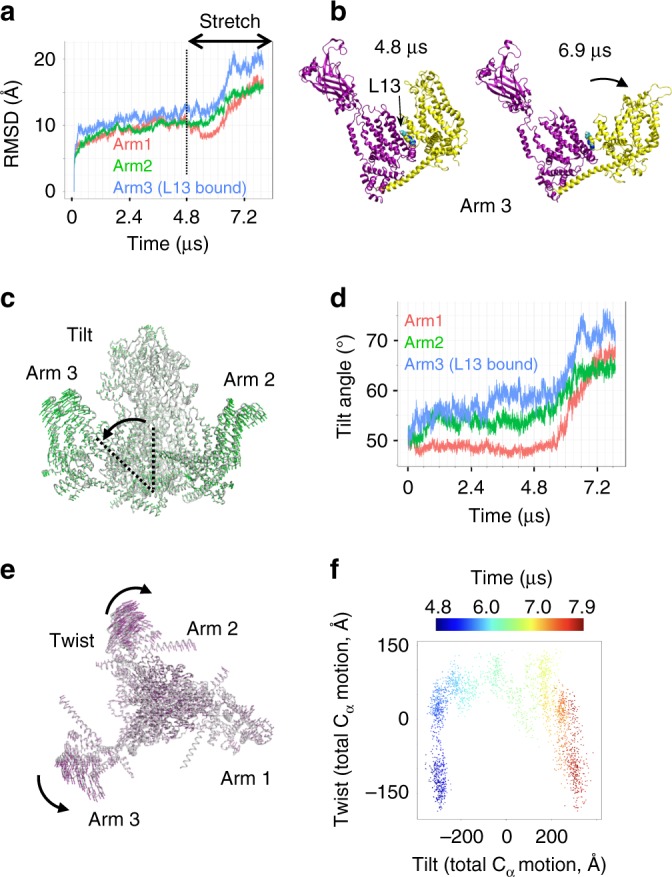
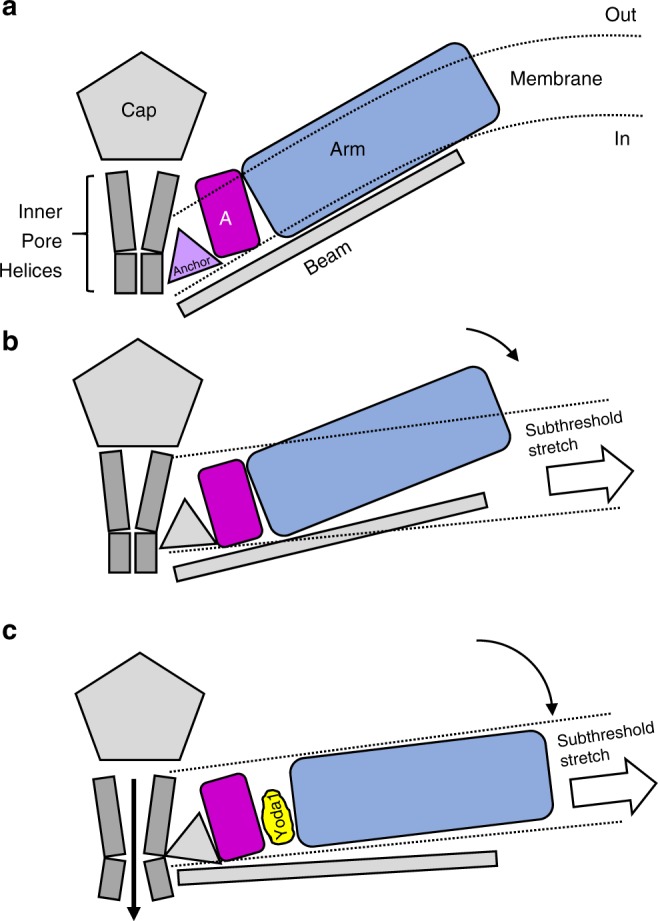
Mechanosensitive Piezo1 and Piezo2 channels transduce various forms of mechanical forces into cellular signals that play vital roles in many important biological processes in vertebrate organisms. Besides mechanical forces, Piezo1 is selectively activated by micromolar concentrations of the small molecule Yoda1 through an unknown mechanism. Here, using a combination of all-atom molecular dynamics simulations, calcium imaging and electrophysiology, we identify an allosteric Yoda1 binding pocket located in the putative mechanosensory domain, approximately 40 Å away from the central pore. Our simulations further indicate that the presence of the agonist correlates with increased tension-induced motions of the Yoda1-bound subunit. Our results suggest a model wherein Yoda1 acts as a molecular wedge, facilitating force-induced conformational changes, effectively lowering the channel's mechanical threshold for activation. The identification of an allosteric agonist binding site in Piezo1 channels will pave the way for the rational design of future Piezo modulators with clinical value.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Wang Fan, Knutson Kaitlyn, Alcaino Constanza, Linden David R., Gibbons Simon J., Kashyap Purna, Grover Madhusudan, Oeckler Richard, Gottlieb Philip A., Li Hui Joyce, Leiter Andrew B., Farrugia Gianrico, Beyder Arthur. Mechanosensitive ion channel Piezo2 is important for enterochromaffin cell response to mechanical forces. The Journal of Physiology. 2016;595(1):79–91. doi: 10.1113/JP272718. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
