

Divergent evolutionary trajectories following speciation in two ectoparasitic honey bee mites
- PMID: 31583288
- PMCID: PMC6773775
- DOI: 10.1038/s42003-019-0606-0
Divergent evolutionary trajectories following speciation in two ectoparasitic honey bee mites
Abstract
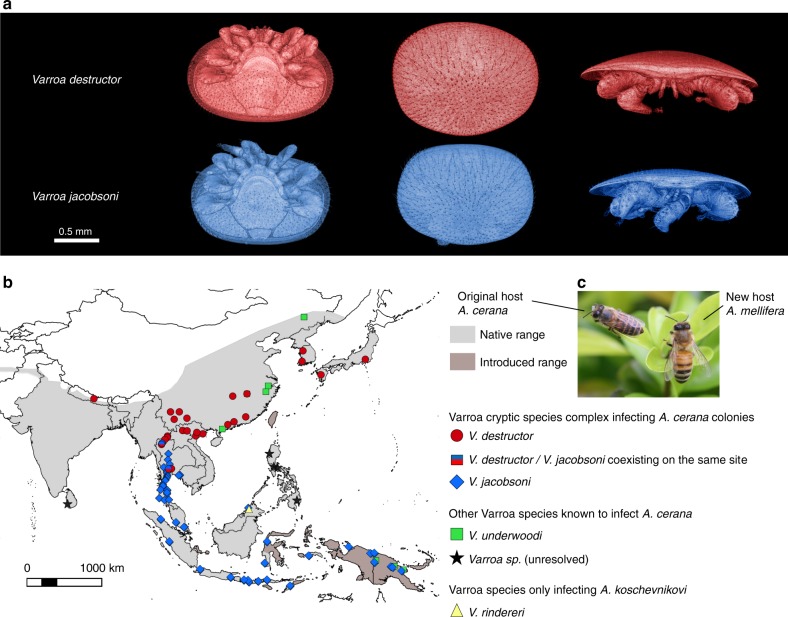
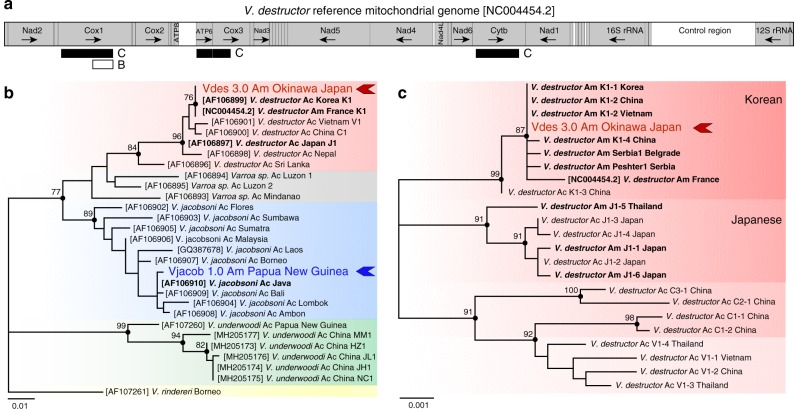
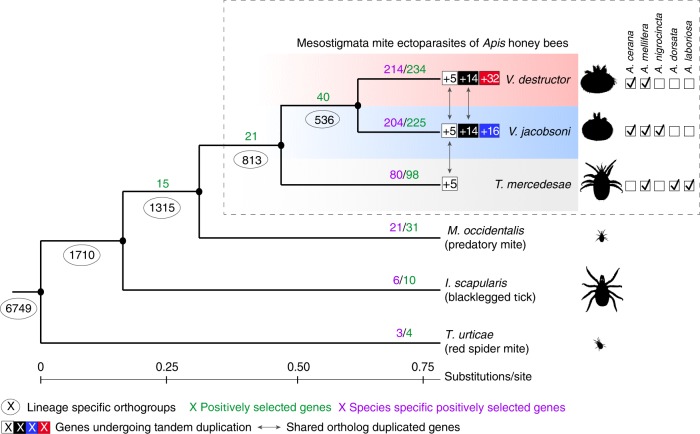
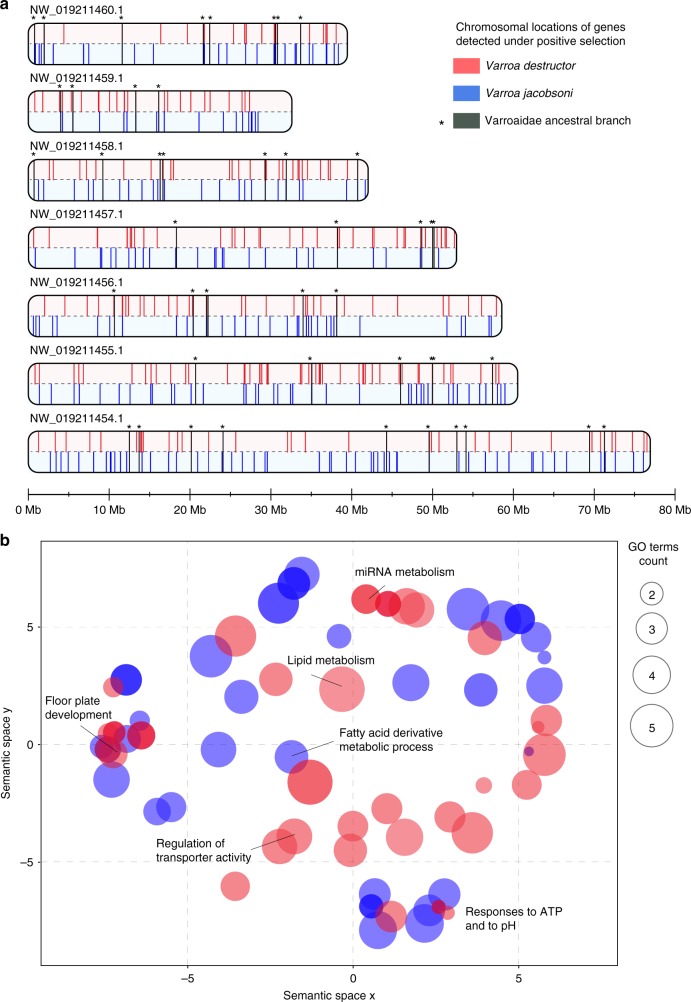
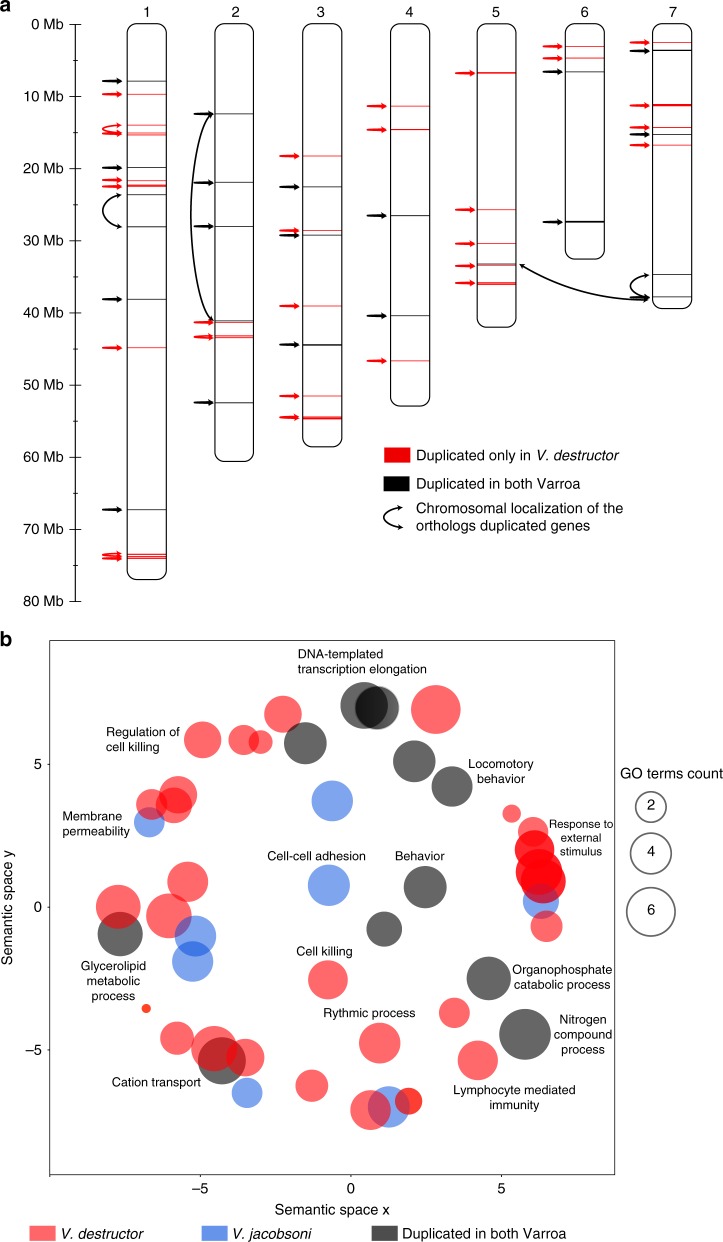
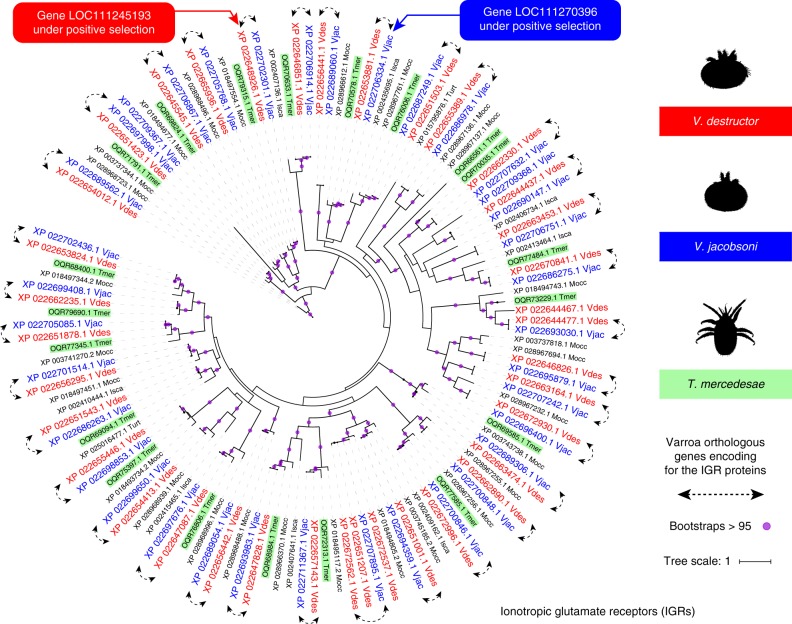
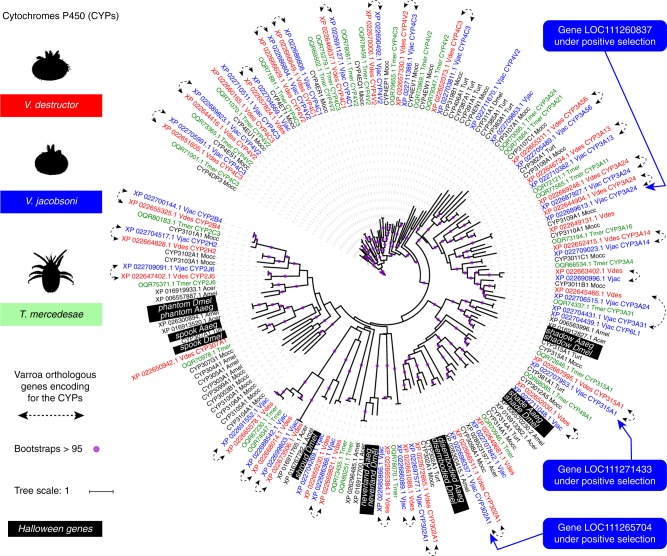
Multispecies host-parasite evolution is common, but how parasites evolve after speciating remains poorly understood. Shared evolutionary history and physiology may propel species along similar evolutionary trajectories whereas pursuing different strategies can reduce competition. We test these scenarios in the economically important association between honey bees and ectoparasitic mites by sequencing the genomes of the sister mite species Varroa destructor and Varroa jacobsoni. These genomes were closely related, with 99.7% sequence identity. Among the 9,628 orthologous genes, 4.8% showed signs of positive selection in at least one species. Divergent selective trajectories were discovered in conserved chemosensory gene families (IGR, SNMP), and Halloween genes (CYP) involved in moulting and reproduction. However, there was little overlap in these gene sets and associated GO terms, indicating different selective regimes operating on each of the parasites. Based on our findings, we suggest that species-specific strategies may be needed to combat evolving parasite communities.
Keywords: Coevolution; Comparative genomics.
© The Author(s) 2019.
Conflict of interest statement
Competing interestsI.L. and S.T.S. declare the following competing interests: I.L. and S.T.S. are employees and shareholders of Phase Genomics, a company commercializing proximity-ligation technology. M.A.T., R.V.R., J.M.K.R., M.L.G., A.K.C., J.D.E., and A.S.M. declare no financial or non-financial competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
