

Metformin Restores CNS Remyelination Capacity by Rejuvenating Aged Stem Cells
- PMID: 31585093
- PMCID: PMC6863391
- DOI: 10.1016/j.stem.2019.08.015
Metformin Restores CNS Remyelination Capacity by Rejuvenating Aged Stem Cells
Abstract
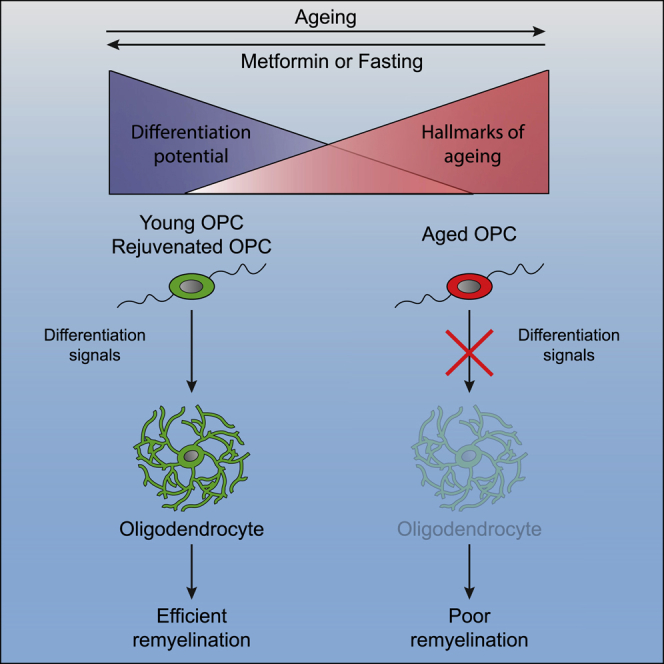
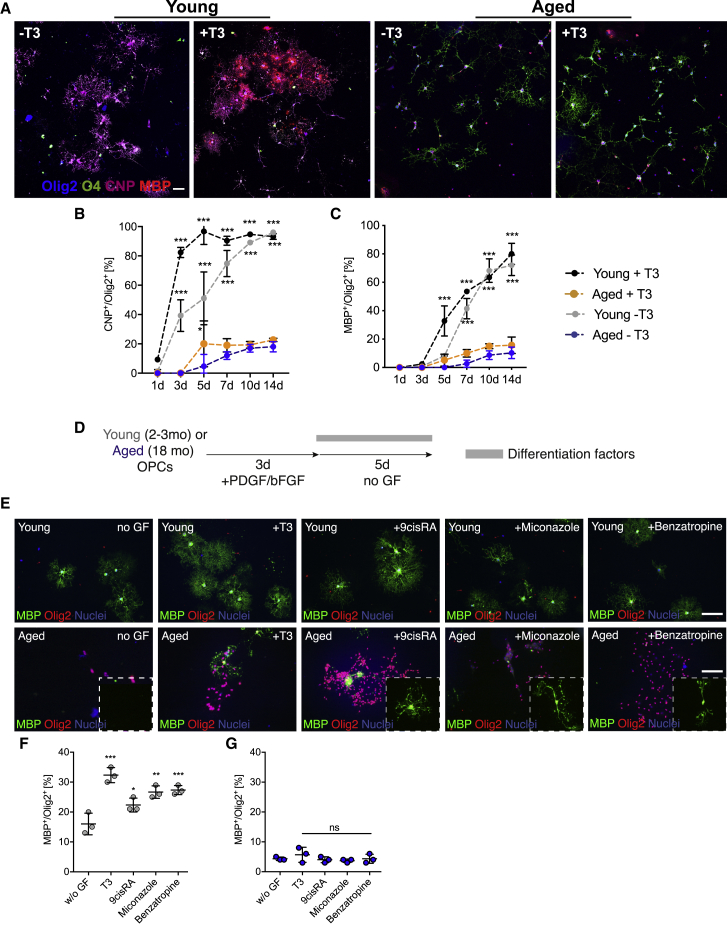
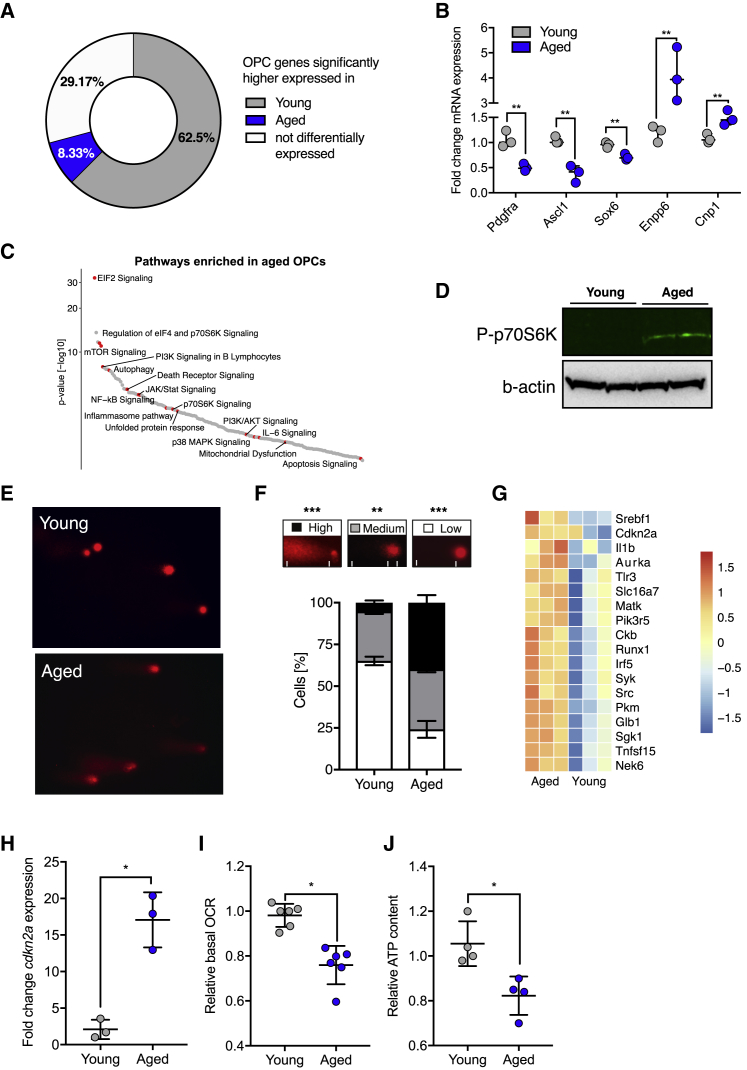
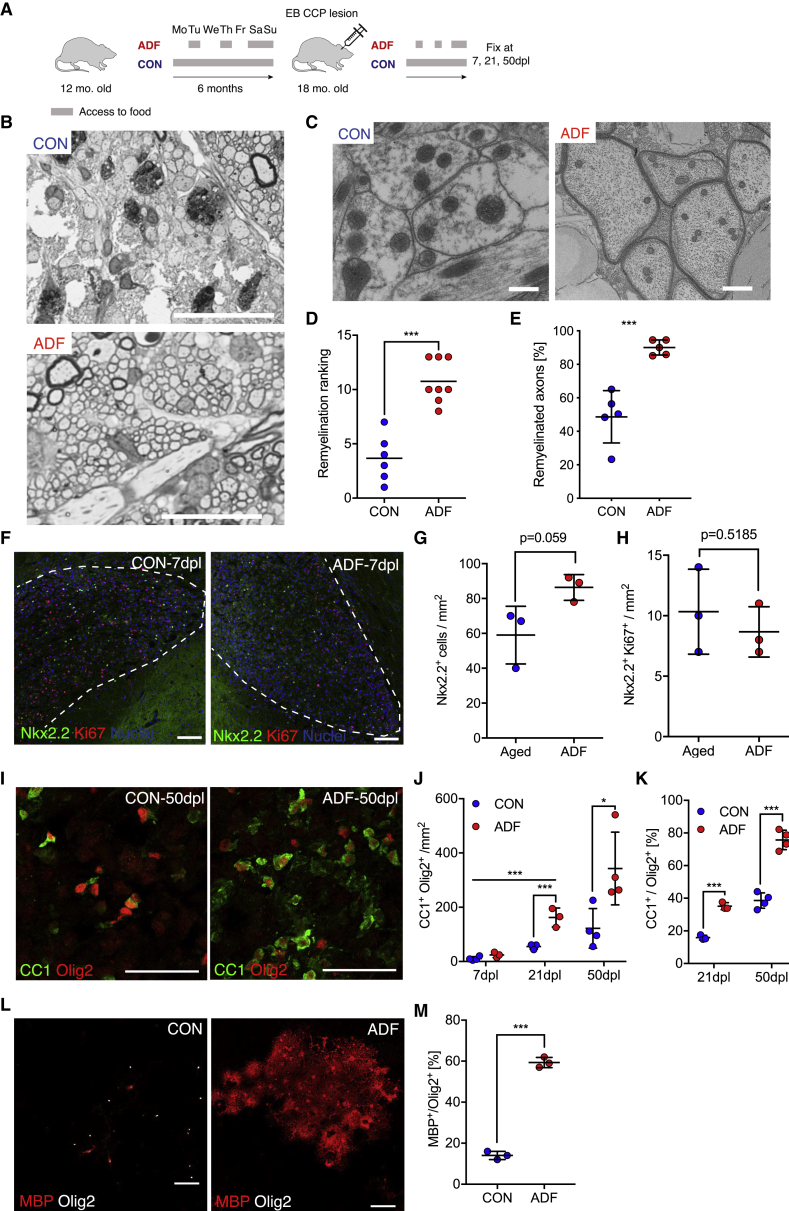
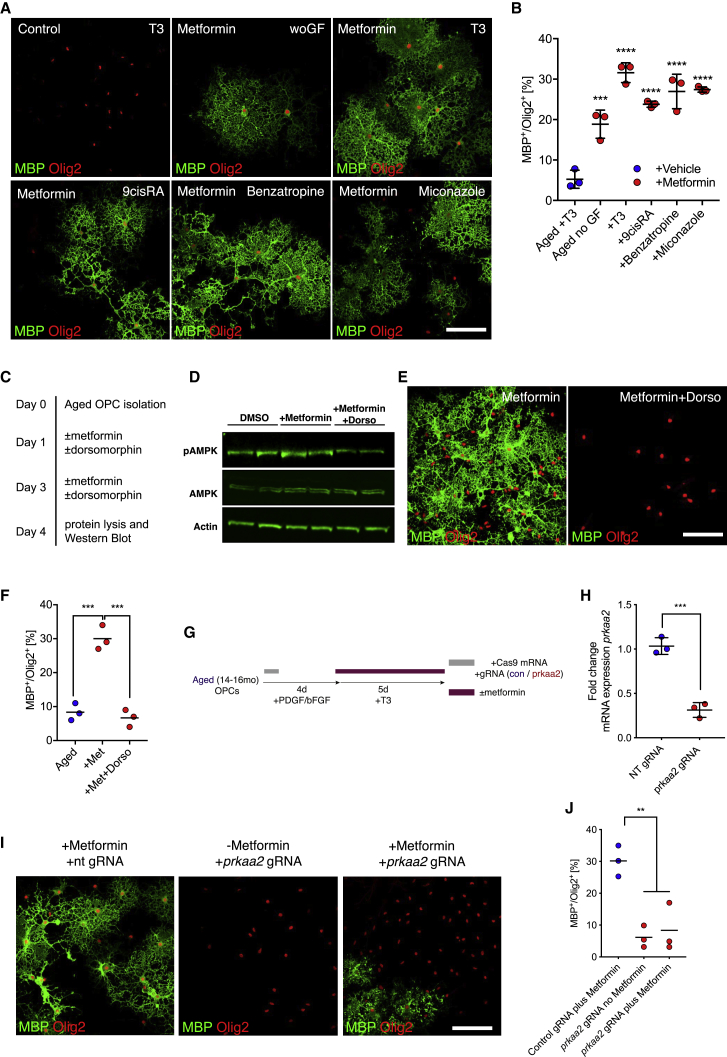
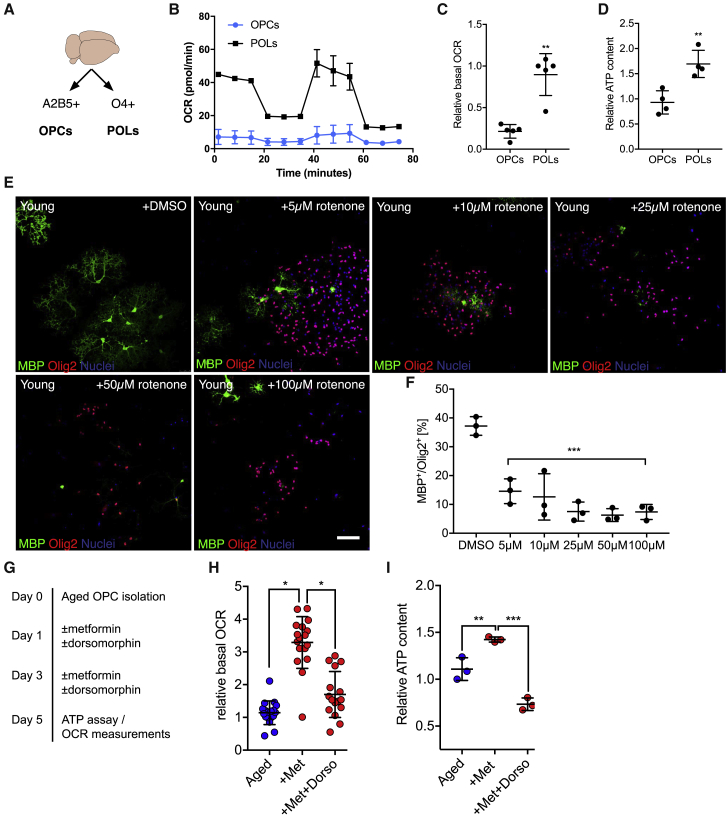
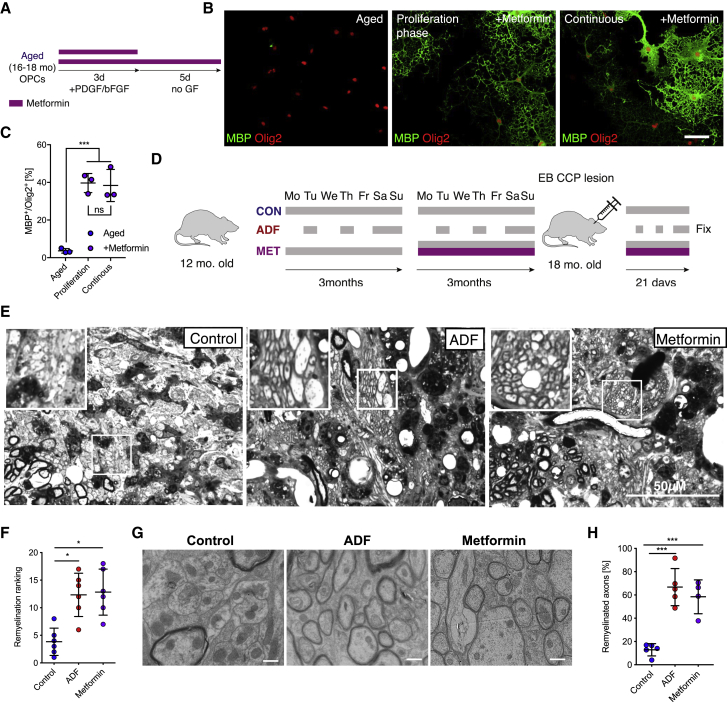
The age-related failure to produce oligodendrocytes from oligodendrocyte progenitor cells (OPCs) is associated with irreversible neurodegeneration in multiple sclerosis (MS). Consequently, regenerative approaches have significant potential for treating chronic demyelinating diseases. Here, we show that the differentiation potential of adult rodent OPCs decreases with age. Aged OPCs become unresponsive to pro-differentiation signals, suggesting intrinsic constraints on therapeutic approaches aimed at enhancing OPC differentiation. This decline in functional capacity is associated with hallmarks of cellular aging, including decreased metabolic function and increased DNA damage. Fasting or treatment with metformin can reverse these changes and restore the regenerative capacity of aged OPCs, improving remyelination in aged animals following focal demyelination. Aged OPCs treated with metformin regain responsiveness to pro-differentiation signals, suggesting synergistic effects of rejuvenation and pro-differentiation therapies. These findings provide insight into aging-associated remyelination failure and suggest therapeutic interventions for reversing such declines in chronic disease.
Keywords: CNS regeneration; adult stem cell; aging; dietary restriction; metformin; oligodendrocyte progenitor cell; rejuvenation; remyelination.
Copyright © 2019 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare that there are patent applications pending related to this work.
Figures
Comment in
-
OPCs on a Diet: A Youthful Serving of Remyelination.Cell Metab. 2019 Dec 3;30(6):1004-1006. doi: 10.1016/j.cmet.2019.11.009. Cell Metab. 2019. PMID: 31801054
References
-
- Blakemore W.F. Pattern of remyelination in the CNS. Nature. 1974;249:577–578. - PubMed
-
- Cantuti-Castelvetri L., Fitzner D., Bosch-Queralt M., Weil M.T., Su M., Sen P., Ruhwedel T., Mitkovski M., Trendelenburg G., Lütjohann D. Defective cholesterol clearance limits remyelination in the aged central nervous system. Science. 2018;359:684–688. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
