

Towards coeliac-safe bread
- PMID: 31585498
- PMCID: PMC7061869
- DOI: 10.1111/pbi.13273
Towards coeliac-safe bread
Abstract
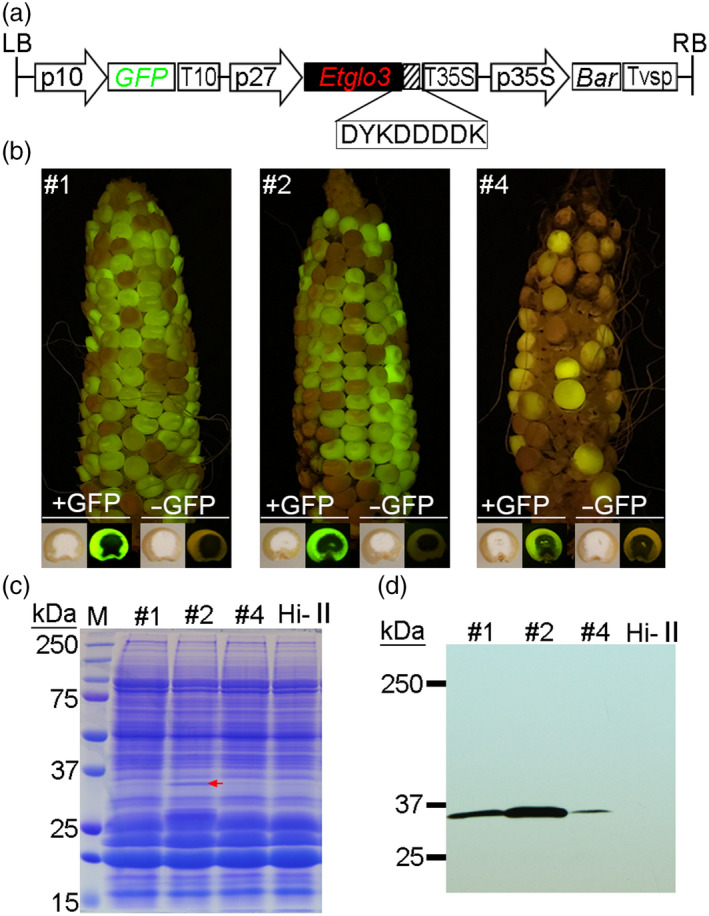
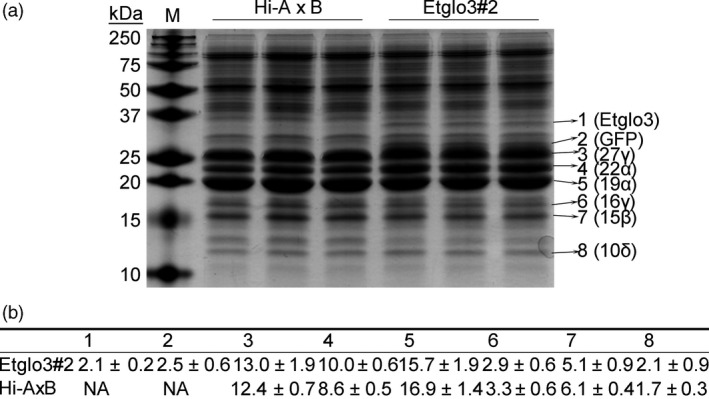
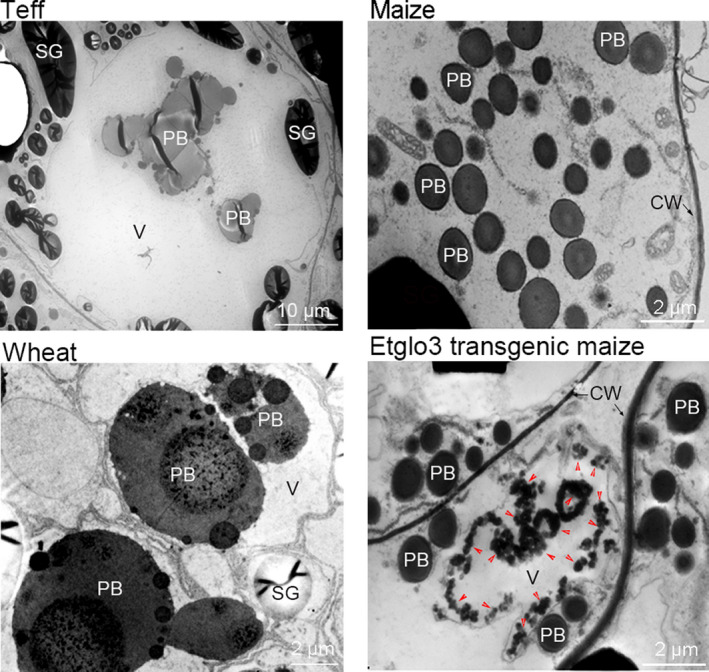
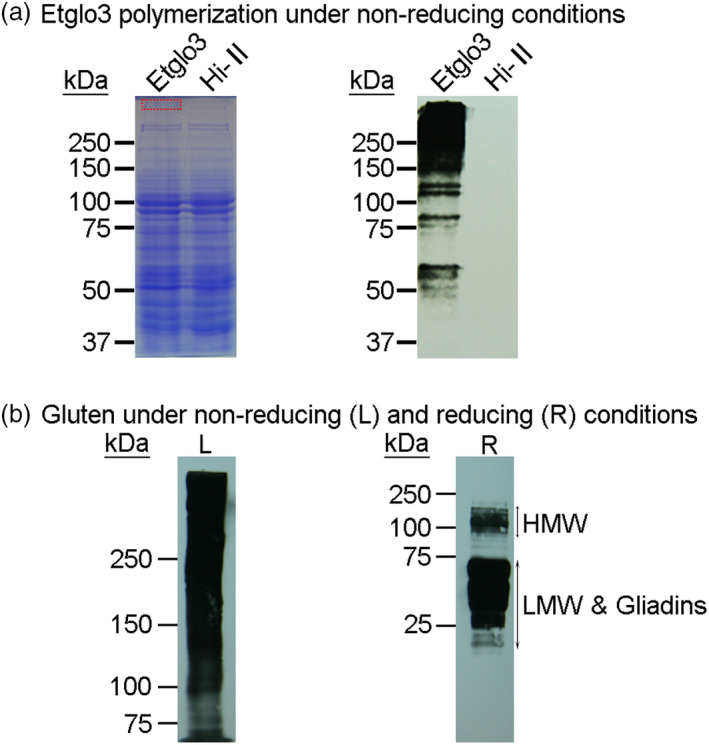
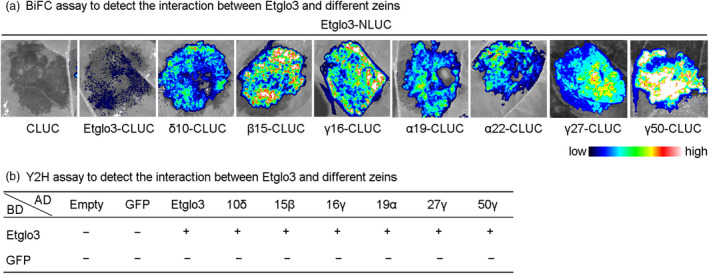
Gluten-free foods cannot substitute for products made from wheat flour. When wheat products are digested, the remaining peptides can trigger an autoimmune disease in 1% of the North American and European population, called coeliac disease. Because wheat proteins are encoded by a large gene family, it has been impossible to use conventional breeding to select wheat varieties that are coeliac-safe. However, one can test the properties of protein variants by expressing single genes in coeliac-safe cereals like maize. One source of protein that can be considered as coeliac-safe and has bread-making properties is teff (Eragrostis tef), a grain consumed in Ethiopia. Here, we show that teff α-globulin3 (Etglo3) forms storage vacuoles in maize that are morphologically similar to those of wheat. Using transmission electron microscopy, immunogold labelling shows that Etglo3 is almost exclusively deposited in the storage vacuole as electron-dense aggregates. Of maize seed storage proteins, 27-kDa γ-zein is co-deposited with Etglo3. Etglo3 polymerizes via intermolecular disulphide bonds in maize, similar to wheat HMW glutenins under non-reducing conditions. Crossing maize Etglo3 transgenic lines with α-, β- and γ-zein RNA interference (RNAi) lines reveals that Etglo3 accumulation is only dramatically reduced in γ-zein RNAi background. This suggests that Etglo3 and 27-kDa γ-zein together cause storage vacuole formation and behave similar to the interactions of glutenins and gliadins in wheat. Therefore, expression of teff α-globulins in maize presents a major step in the development of a coeliac-safe grain with bread-making properties.
Keywords: gluten; maize; storage vacuole; teff; wheat; α-globulin.
© 2019 The Authors. Plant Biotechnology Journal published by Society for Experimental Biology and The Association of Applied Biologists and John Wiley & Sons Ltd.
Conflict of interest statement
The authors have no conflict of interest to declare.
Figures


Similar articles
-
Targeting of prolamins by RNAi in bread wheat: effectiveness of seven silencing-fragment combinations for obtaining lines devoid of coeliac disease epitopes from highly immunogenic gliadins.Plant Biotechnol J. 2016 Mar;14(3):986-96. doi: 10.1111/pbi.12455. Epub 2015 Aug 24. Plant Biotechnol J. 2016. PMID: 26300126 Free PMC article.
-
Outlook for coeliac disease patients: towards bread wheat with hypoimmunogenic gluten by gene editing of α- and γ-gliadin gene families.BMC Plant Biol. 2019 Aug 1;19(1):333. doi: 10.1186/s12870-019-1889-5. BMC Plant Biol. 2019. PMID: 31370789 Free PMC article.
-
Interaction of maize zein with wheat gluten in composite dough and bread as determined by confocal laser scanning microscopy.Scanning. 2002 Jan-Feb;24(1):1-5. doi: 10.1002/sca.4950240101. Scanning. 2002. PMID: 11866341
-
Chemical composition and food uses of teff (Eragrostis tef).Food Chem. 2018 Jan 15;239:402-415. doi: 10.1016/j.foodchem.2017.06.101. Epub 2017 Jun 20. Food Chem. 2018. PMID: 28873585 Review.
-
Gluten Free Wheat: Are We There?Nutrients. 2019 Feb 26;11(3):487. doi: 10.3390/nu11030487. Nutrients. 2019. PMID: 30813572 Free PMC article. Review.
Cited by
-
Gluten Unraveled: Latest Insights on Terminology, Diagnosis, Pathophysiology, Dietary Strategies, and Intestinal Microbiota Modulations-A Decade in Review.Nutrients. 2024 Oct 25;16(21):3636. doi: 10.3390/nu16213636. Nutrients. 2024. PMID: 39519469 Free PMC article. Review.
References
-
- Adebowale, A.R.A. , Emmambux, M.N. , Beukes, M. and Taylor, J.R.N. (2011) Fractionation and characterization of teff proteins. J. Cereal Sci. 54, 380–386.
-
- Bechtel, D.B. and Juliano, B.O. (1980) Formation of protein bodies in the starchy endosperm of rice (Oryza‐Sativa‐L) – a reinvestigation. Ann. Bot. 45, 503–509.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
