

The protective role of microRNA-21 against coxsackievirus B3 infection through targeting the MAP2K3/P38 MAPK signaling pathway
- PMID: 31585536
- PMCID: PMC6778380
- DOI: 10.1186/s12967-019-2077-y
The protective role of microRNA-21 against coxsackievirus B3 infection through targeting the MAP2K3/P38 MAPK signaling pathway
Abstract
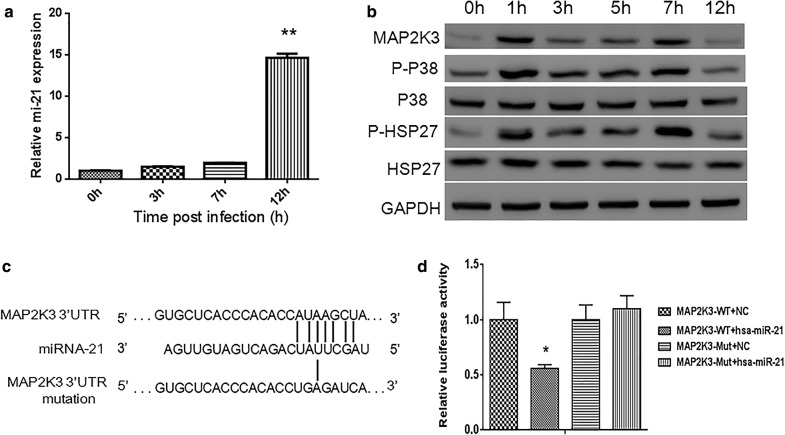
Background: The P38 mitogen-activated protein kinase (MAPK) pathway plays an essential role in CVB3-induced diseases. We previously demonstrated microRNA-21 has potential inhibitory effect on the MAP2K3 which locates upstream of P38 MAPK and was upregulated in mouse hearts upon CVB3 infection. However, the effect and underlying mechanism of miRNA-21 on CVB3 infection remain unclear.
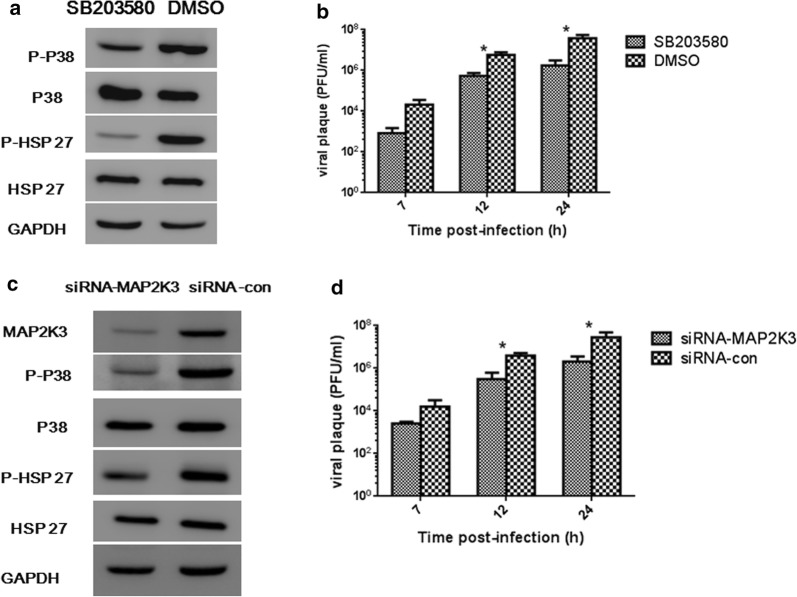
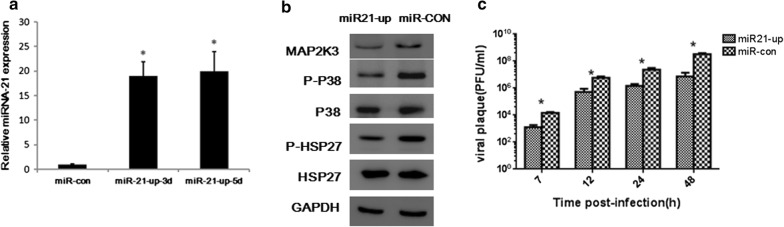
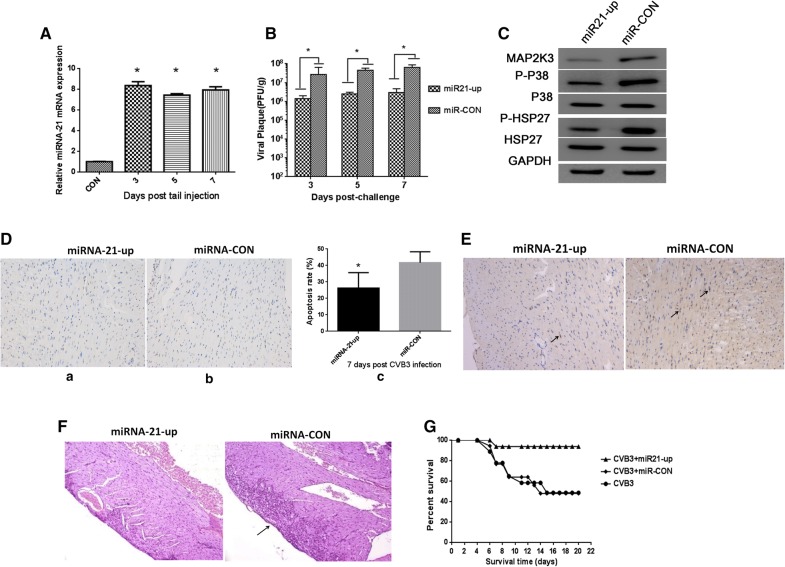
Methods: We detected continuous changes of cellular miRNA-21 and P38 MAPK proteins expression profiling post CVB3 infection in vitro within 12 h. P38 MAPK signaling was inhibited by the specific inhibitor, small interfering RNA and miRNA-21 mimic in vitro, CVB3 replication, cell apoptosis rate and proliferation were detected. Viral load in the mice heart, cardiomyocyte apoptosis rate and histological of the heart were also detected in the mice model of viral myocarditis pretreated with miRNA-21-lentivirus.
Results: We observed significant upregulation of miRNA-21 expression followed by suppression of the MAP2K3/P38 MAPK signaling in CVB3-infected Hela cells. The inactivation of the MAP2K3/P38 MAPK signaling by P38 MAPK specific inhibitor, small interfering RNA against MAP2K3, or miRNA-21 overexpression significantly inhibited viral progeny release from CVB3-infected cells. Mechanistically, when compared with control miRNA, miRNA-21 showed no effect on capsid protein VP1 expression and viral load within host cells, while significantly reversing CVB3-induced caspase-3 activation and cell apoptosis rate, further promoting proliferation of infected cells, which indicates the inhibitory effect of miRNA-21 on CVB3 progeny release. In the in vivo study, when compared with control miRNA, miRNA-21 pretreatment remarkably inactivated the MAP2K3/P38 MAPK signaling in mice and protected them against CVB3 infection as evidenced by significantly alleviated cell apoptosis rate, reduced viral titers, necrosis in the heart as well as by remarkably prolonged survival time.
Conclusions: miRNA-21 were reverse correlated with P38 MAPK activation post CVB3 infection, miRNA-21 overexpression significantly inhibited viral progeny release and decreased myocytes apoptosis rate in vitro and in vivo, suggesting that miRNA-21 may serve as a potential therapeutic agent against CVB3 infection through targeting the MAP2K3/P38 MAPK signaling.
Keywords: CVB3; P38 MAPK; Viral release; miRNA-21.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

Similar articles
-
The lncRNA MEG3/miRNA-21/P38MAPK axis inhibits coxsackievirus 3 replication in acute viral myocarditis.Virus Res. 2024 Jan 2;339:199250. doi: 10.1016/j.virusres.2023.199250. Epub 2023 Nov 1. Virus Res. 2024. PMID: 37865350 Free PMC article.
-
Stress-activated protein kinases are involved in coxsackievirus B3 viral progeny release.J Virol. 2005 Nov;79(22):13875-81. doi: 10.1128/JVI.79.22.13875-13881.2005. J Virol. 2005. PMID: 16254323 Free PMC article.
-
Coxsackievirus B3 replication is reduced by inhibition of the extracellular signal-regulated kinase (ERK) signaling pathway.J Virol. 2002 Apr;76(7):3365-73. doi: 10.1128/jvi.76.7.3365-3373.2002. J Virol. 2002. PMID: 11884562 Free PMC article.
-
A Kidnapping Story: How Coxsackievirus B3 and Its Host Cell Interact.Cell Physiol Biochem. 2019;53(1):121-140. doi: 10.33594/000000125. Cell Physiol Biochem. 2019. PMID: 31230428 Review.
-
p38 activation and viral infection.Expert Rev Mol Med. 2022 Jan 21;24:e4. doi: 10.1017/erm.2021.29. Expert Rev Mol Med. 2022. PMID: 35060846 Review.
Cited by
-
The Role of Non-coding RNAs in Viral Myocarditis.Front Cell Infect Microbiol. 2020 Jul 2;10:312. doi: 10.3389/fcimb.2020.00312. eCollection 2020. Front Cell Infect Microbiol. 2020. PMID: 32754448 Free PMC article. Review.
-
MicroRNA-22-3p displaces critical host factors from the 5' UTR and inhibits the translation of Coxsackievirus B3 RNA.J Virol. 2024 Feb 20;98(2):e0150423. doi: 10.1128/jvi.01504-23. Epub 2024 Jan 30. J Virol. 2024. PMID: 38289119 Free PMC article.
-
Comprehensive Analysis of Differences in N6-Methyladenosine RNA Methylation Groups in CVB3-Induced Viral Myocarditis and Identification of the Anti-Apoptotic Role of RBM15B.J Inflamm Res. 2025 Jun 17;18:7933-7949. doi: 10.2147/JIR.S503823. eCollection 2025. J Inflamm Res. 2025. PMID: 40546402 Free PMC article.
-
Ly6G+ Neutrophils and Interleukin-17 Are Essential in Protection against Rodent Malaria Caused by Plasmodium berghei ANKA.Research (Wash D C). 2024 Dec 19;7:0559. doi: 10.34133/research.0559. eCollection 2024. Research (Wash D C). 2024. PMID: 39703777 Free PMC article.
-
Role of non-coding RNAs in the pathogenesis of viral myocarditis.Virulence. 2025 Dec;16(1):2466480. doi: 10.1080/21505594.2025.2466480. Epub 2025 Feb 20. Virulence. 2025. PMID: 39950847 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
