

Differential Modulation of Human Innate Lymphoid Cell (ILC) Subsets by IL-10 and TGF-β
- PMID: 31586075
- PMCID: PMC6778123
- DOI: 10.1038/s41598-019-50308-8
Differential Modulation of Human Innate Lymphoid Cell (ILC) Subsets by IL-10 and TGF-β
Abstract
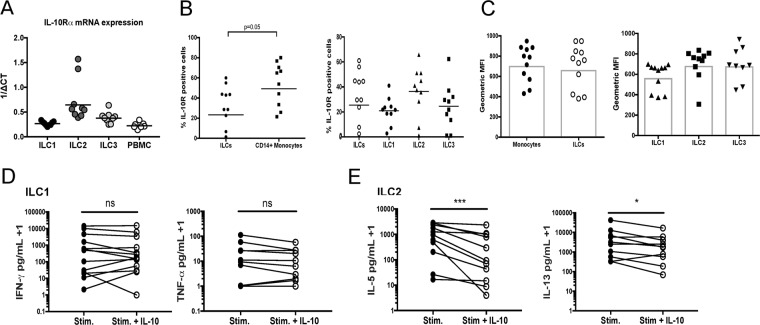
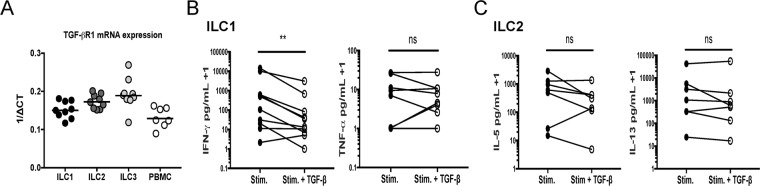
Using multiparameter flow cytometry human innate lymphoid cell (ILC) subsets can be detected in the circulation, in relatively low frequencies. Despite the low frequency of ILCs in circulation, ex vivo experiments have demonstrated that these ILCs release extremely large per cell quantities of signature ILC cytokines following activation. To determine how activated ILC cytokine production is regulated, ILC subsets were activated in the presence or absence of the immunoregulatory cytokines IL-10 and TGF-β. An examination of circulating ILC subsets revealed surface expression of IL-10Rα and mRNA expression of both IL-10Rα and TGF-βR1 for all ILC subsets. Stimulated ILC1 production of IFN-γ was decreased by TGF-β and not IL-10. Interestingly, ILC2s stimulated in the presence of IL-10 had a marked reduction in cytokine production of IL-5 and IL-13 while TGF-β had no effect on ILC2 cytokine production. Ex vivo activated ILC1 and ILC2 subsets were also found to be a source of the immunoregulatory cytokine IL-10, raising the potential for ILC-mediated regulation of immune cells. These findings demonstrate the differential effects of immunoregulatory cytokines IL-10 and TGF-β on activated ILC1 and ILC2 populations ex vivo.
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
Type 2 Diabetes Mellitus and Latent Tuberculosis Infection Moderately Influence Innate Lymphoid Cell Immune Responses in Uganda.Front Immunol. 2021 Aug 27;12:716819. doi: 10.3389/fimmu.2021.716819. eCollection 2021. Front Immunol. 2021. PMID: 34512639 Free PMC article.
-
Inflammatory status might direct ILC and NK cells to IL-17 expressing ILC3 and NK subsets in Behcet's disease.Immunol Lett. 2021 Jul;235:1-8. doi: 10.1016/j.imlet.2021.04.008. Epub 2021 Apr 23. Immunol Lett. 2021. PMID: 33901541
-
Fingolimod Alters Tissue Distribution and Cytokine Production of Human and Murine Innate Lymphoid Cells.Front Immunol. 2019 Feb 13;10:217. doi: 10.3389/fimmu.2019.00217. eCollection 2019. Front Immunol. 2019. PMID: 30828332 Free PMC article.
-
The Yin and Yang of Innate Lymphoid Cells in Cancer.Immunol Lett. 2016 Nov;179:29-35. doi: 10.1016/j.imlet.2016.06.003. Epub 2016 Jun 11. Immunol Lett. 2016. PMID: 27296768 Review.
-
Dissecting human ILC heterogeneity: more than just three subsets.Immunology. 2018 Mar;153(3):297-303. doi: 10.1111/imm.12862. Epub 2017 Dec 26. Immunology. 2018. PMID: 29140572 Free PMC article. Review.
Cited by
-
Regulatory ILC2-Role of IL-10 Producing ILC2 in Asthma.Cells. 2023 Oct 31;12(21):2556. doi: 10.3390/cells12212556. Cells. 2023. PMID: 37947634 Free PMC article. Review.
-
Regulatory T Cells, a Viable Target Against Airway Allergic Inflammatory Responses in Asthma.Front Immunol. 2022 Jun 10;13:902318. doi: 10.3389/fimmu.2022.902318. eCollection 2022. Front Immunol. 2022. PMID: 35757774 Free PMC article. Review.
-
Helper Innate Lymphoid Cells-Unappreciated Players in Melanoma Therapy.Cancers (Basel). 2023 Feb 1;15(3):933. doi: 10.3390/cancers15030933. Cancers (Basel). 2023. PMID: 36765891 Free PMC article. Review.
-
Recruitment of CXCR4+ type 1 innate lymphoid cells distinguishes sarcoidosis from other skin granulomatous diseases.J Clin Invest. 2024 Sep 3;134(17):e178711. doi: 10.1172/JCI178711. J Clin Invest. 2024. PMID: 39225100 Free PMC article.
-
Dendritic cells mediated by small extracellular vesicles derived from MSCs attenuated the ILC2 activity via PGE2 in patients with allergic rhinitis.Stem Cell Res Ther. 2023 Jul 24;14(1):180. doi: 10.1186/s13287-023-03408-2. Stem Cell Res Ther. 2023. PMID: 37488601 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
