

Intermittent compressive force promotes osteogenic differentiation in human periodontal ligament cells by regulating the transforming growth factor-β pathway
- PMID: 31591384
- PMCID: PMC6779887
- DOI: 10.1038/s41419-019-1992-4
Intermittent compressive force promotes osteogenic differentiation in human periodontal ligament cells by regulating the transforming growth factor-β pathway
Abstract
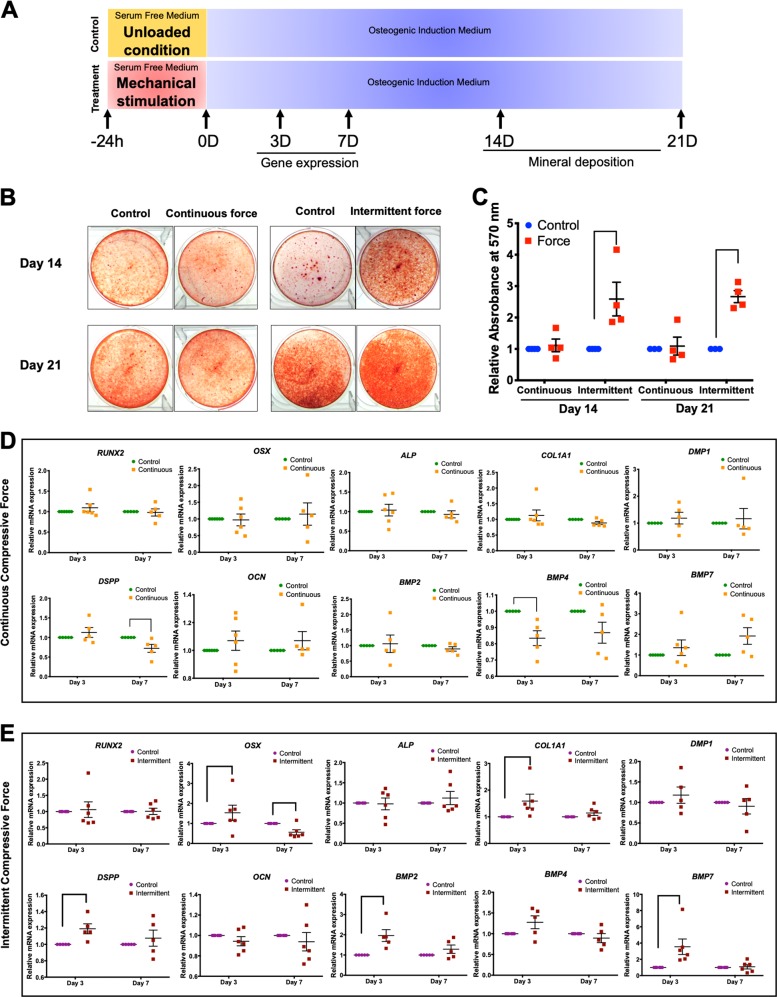
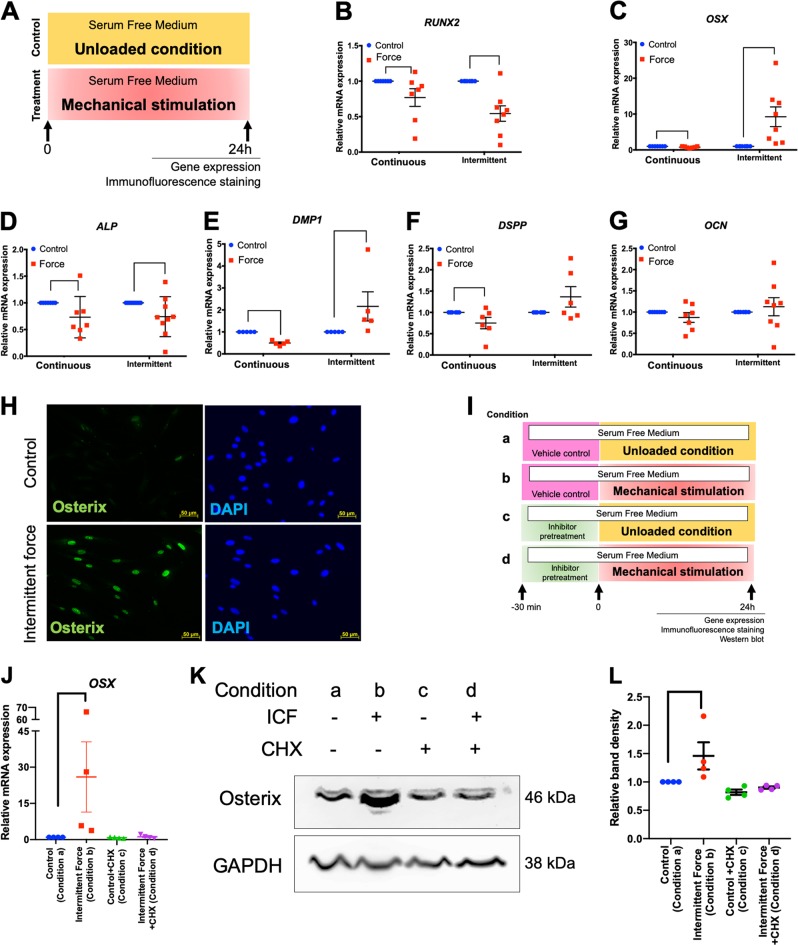
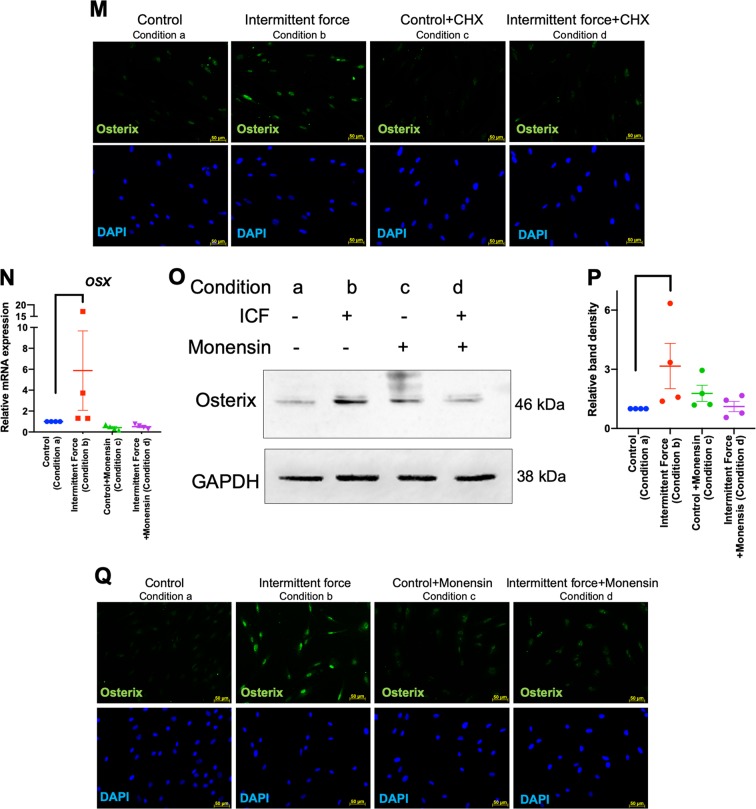
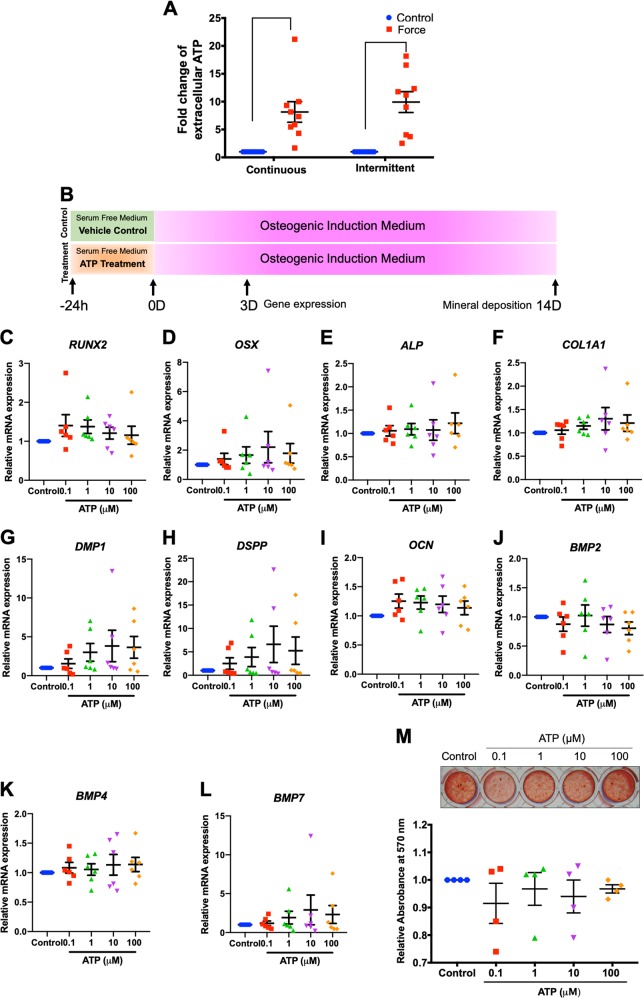
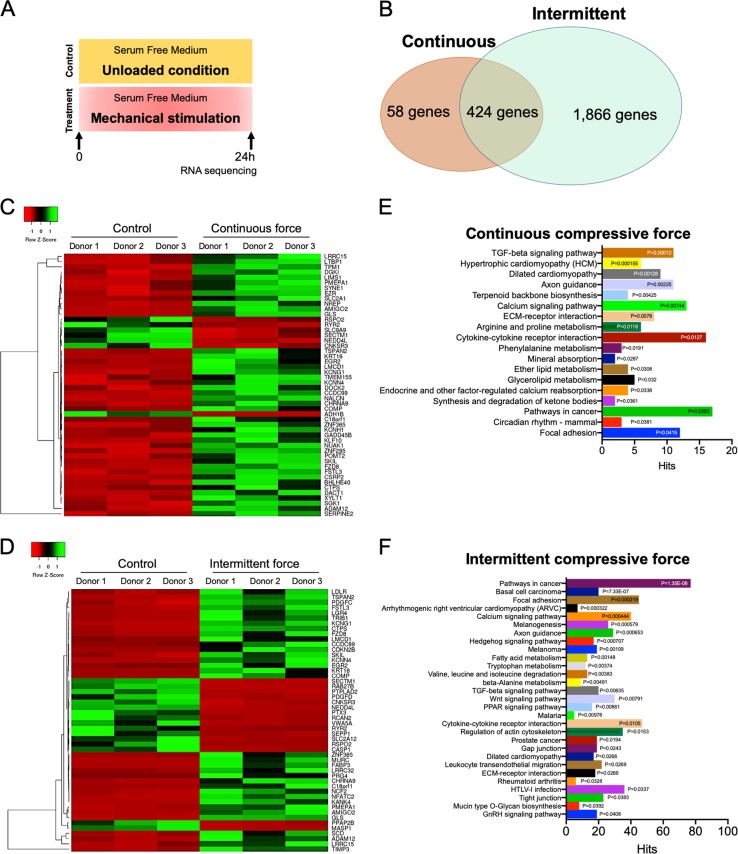
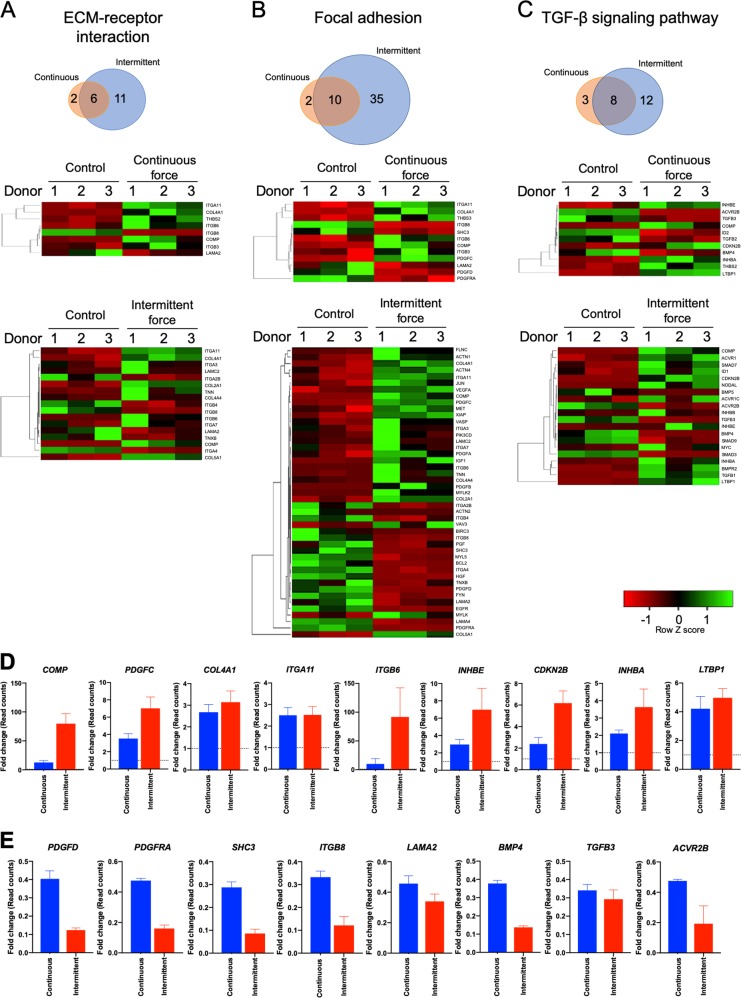
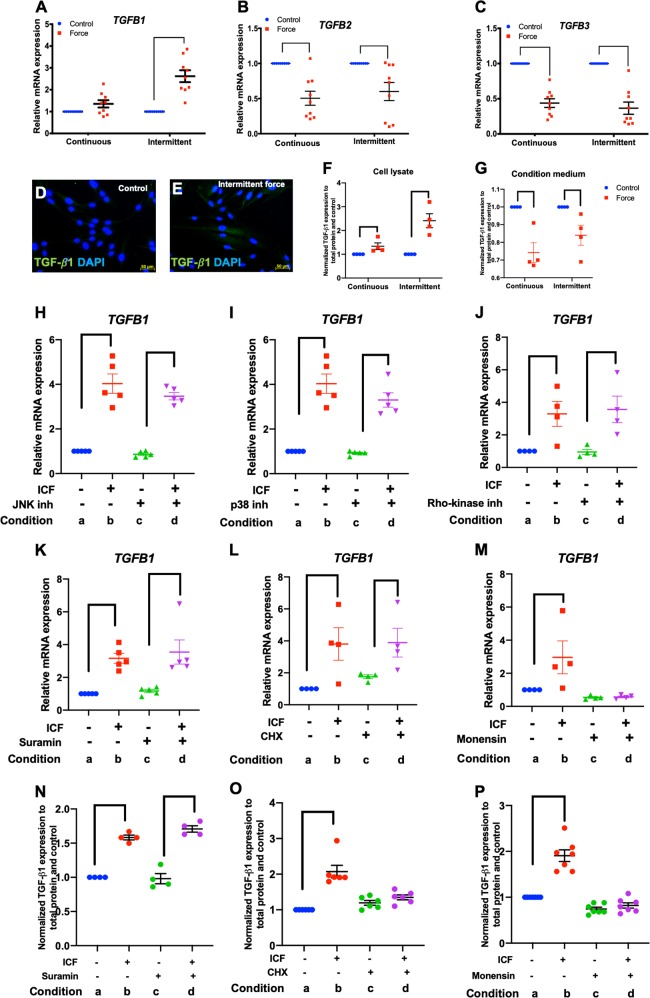
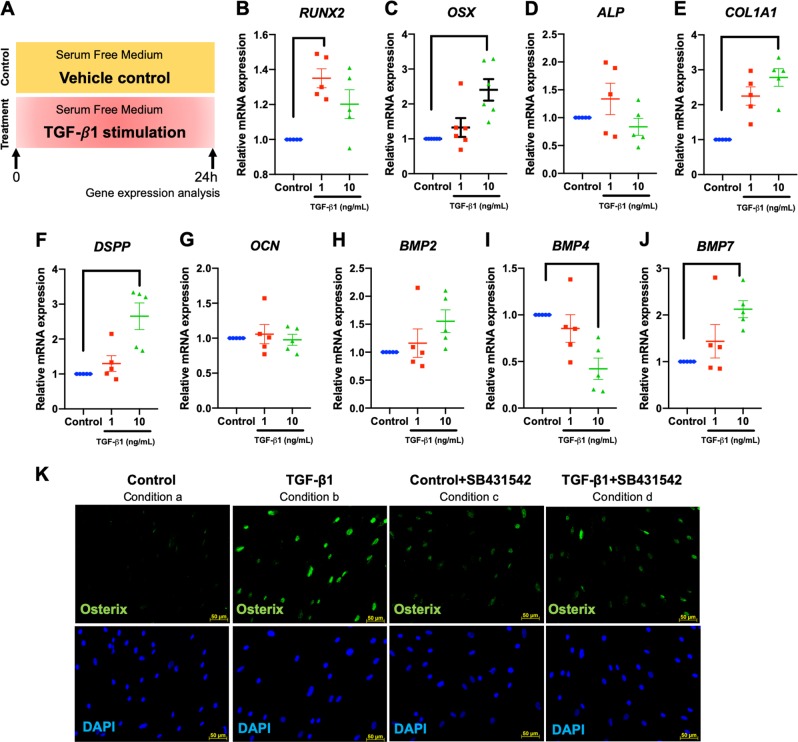
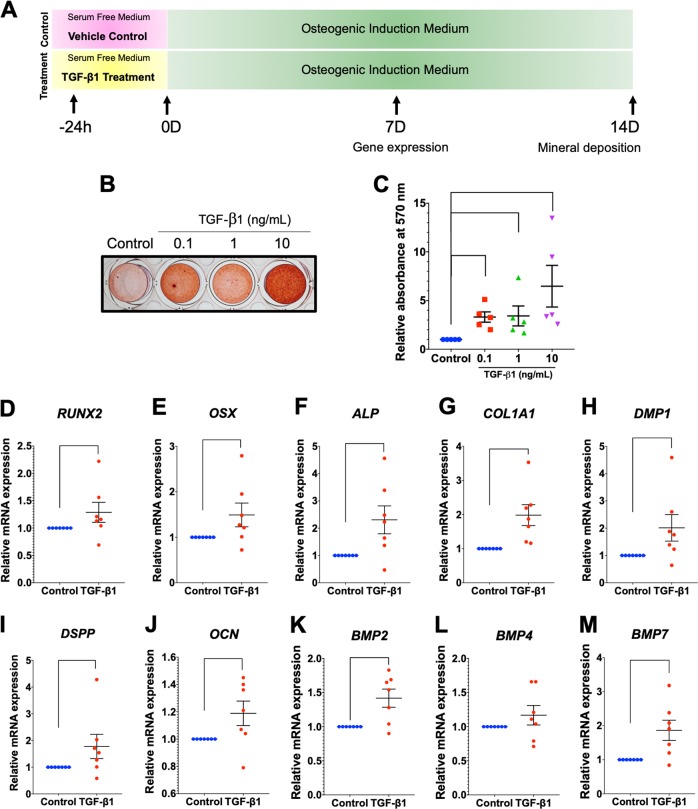
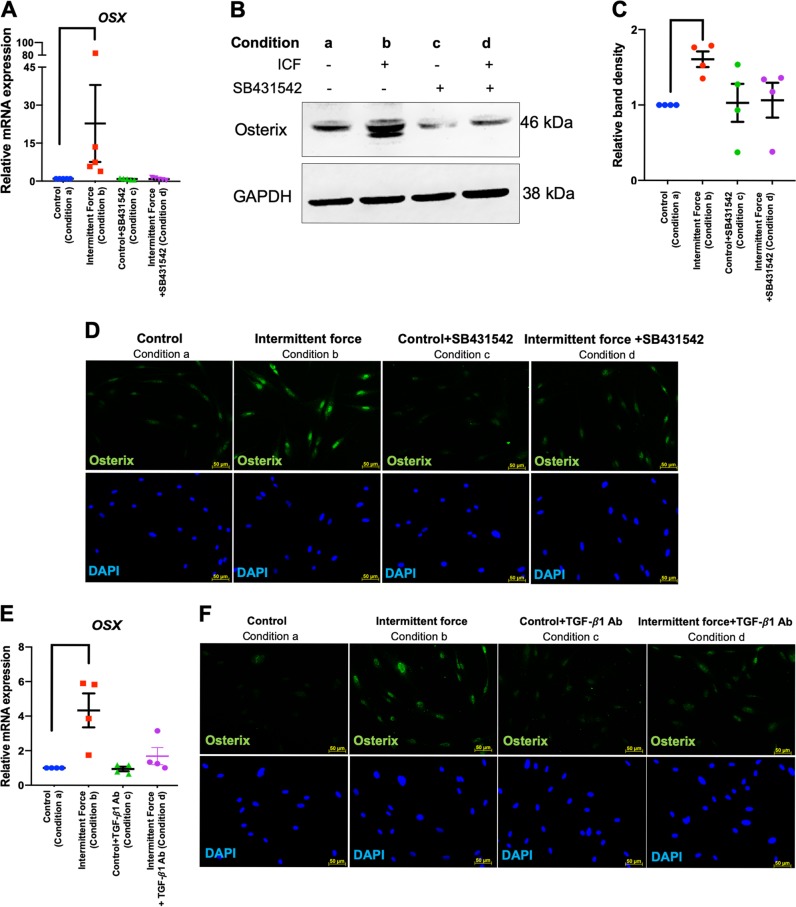
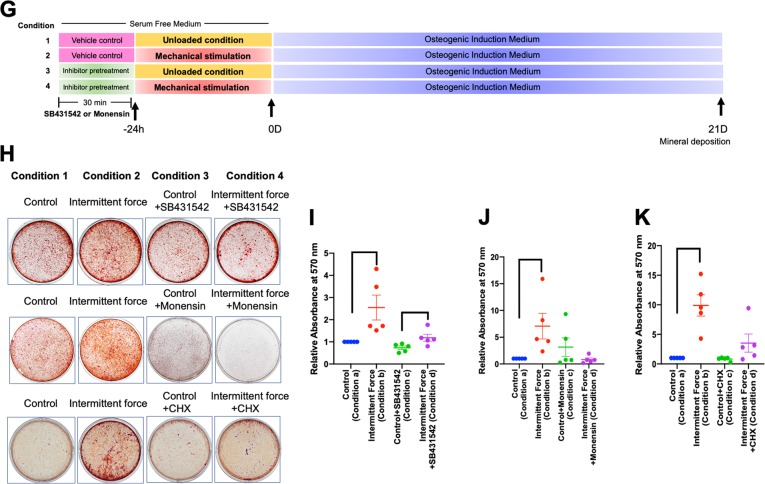
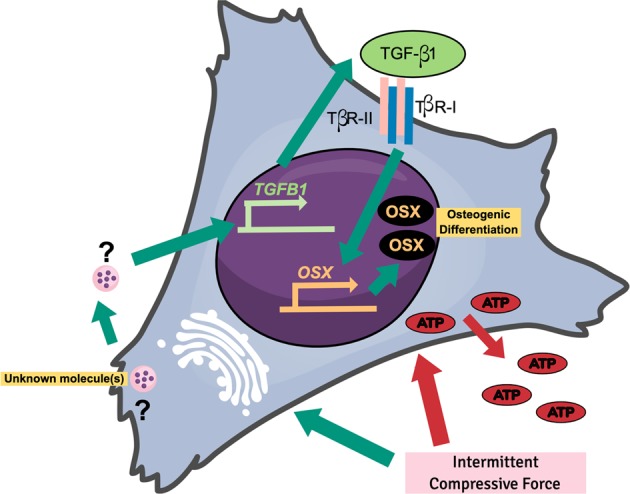
Mechanical force regulates periodontal ligament cell (PDL) behavior. However, different force types lead to distinct PDL responses. Here, we report that pretreatment with an intermittent compressive force (ICF), but not a continuous compressive force (CCF), promoted human PDL (hPDL) osteogenic differentiation as determined by osteogenic marker gene expression and mineral deposition in vitro. ICF-induced osterix (OSX) expression was inhibited by cycloheximide and monensin. Although CCF and ICF significantly increased extracellular adenosine triphosphate (ATP) levels, pretreatment with exogenous ATP did not affect hPDL osteogenic differentiation. Gene-expression profiling of hPDLs subjected to CCF or ICF revealed that extracellular matrix (ECM)-receptor interaction, focal adhesion, and transforming growth factor beta (TGF-β) signaling pathway genes were commonly upregulated, while calcium signaling pathway genes were downregulated in both CCF- and ICF-treated hPDLs. The TGFB1 mRNA level was significantly increased, while those of TGFB2 and TGFB3 were decreased by ICF treatment. In contrast, CCF did not modify TGFB1 expression. Inhibiting TGF-β receptor type I or adding a TGF-β1 neutralizing antibody attenuated the ICF-induced OSX expression. Exogenous TGF-β1 pretreatment promoted hPDL osteogenic marker gene expression and mineral deposition. Additionally, pretreatment with ICF in the presence of TGF-β receptor type I inhibitor attenuated the ICF-induced mineralization. In conclusion, this study reveals the effects of ICF on osteogenic differentiation in hPDLs and implicates TGF-β signaling as one of its regulatory mechanisms.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
