

Rapid selection and identification of functional CD8+ T cell epitopes from large peptide-coding libraries
- PMID: 31591401
- PMCID: PMC6779888
- DOI: 10.1038/s41467-019-12444-7
Rapid selection and identification of functional CD8+ T cell epitopes from large peptide-coding libraries
Abstract
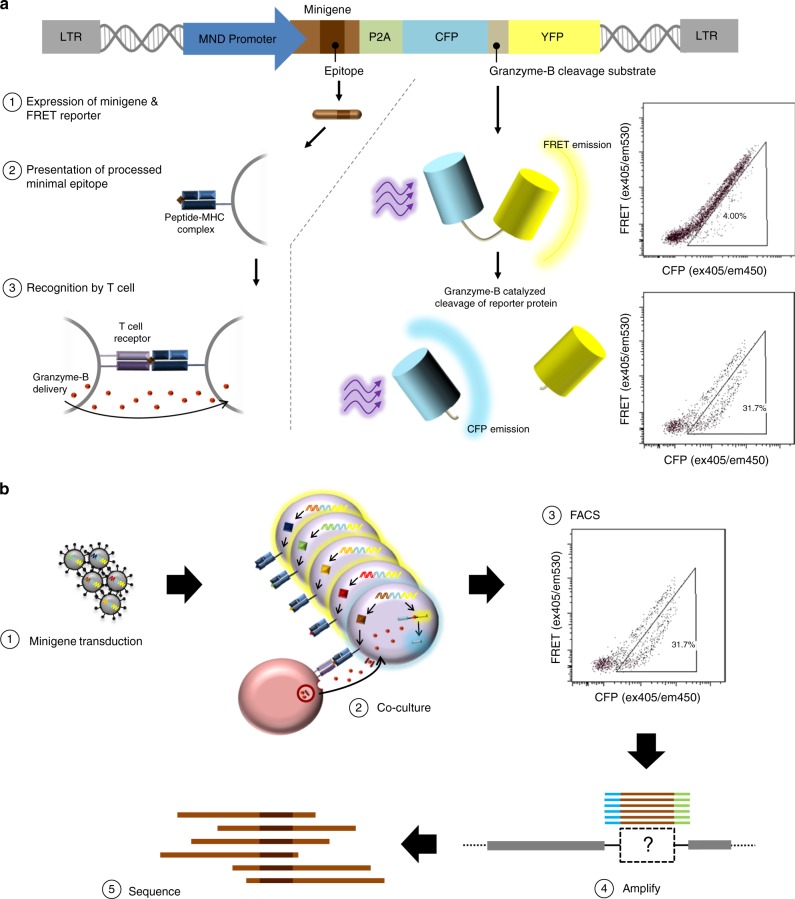
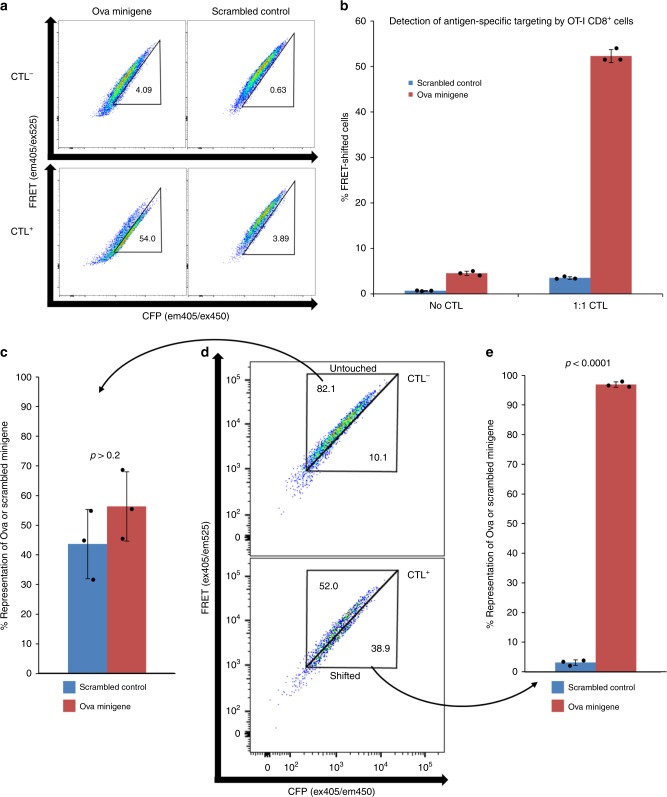
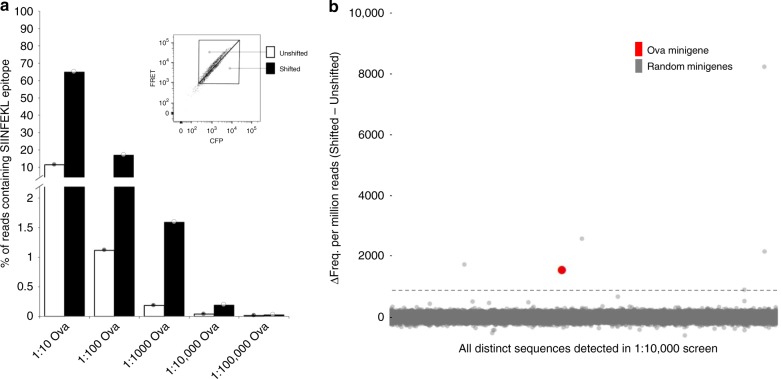
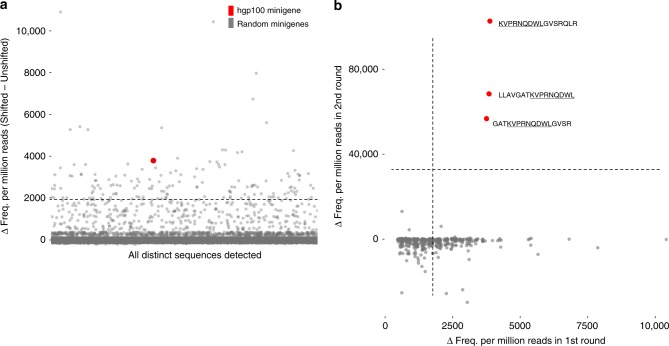
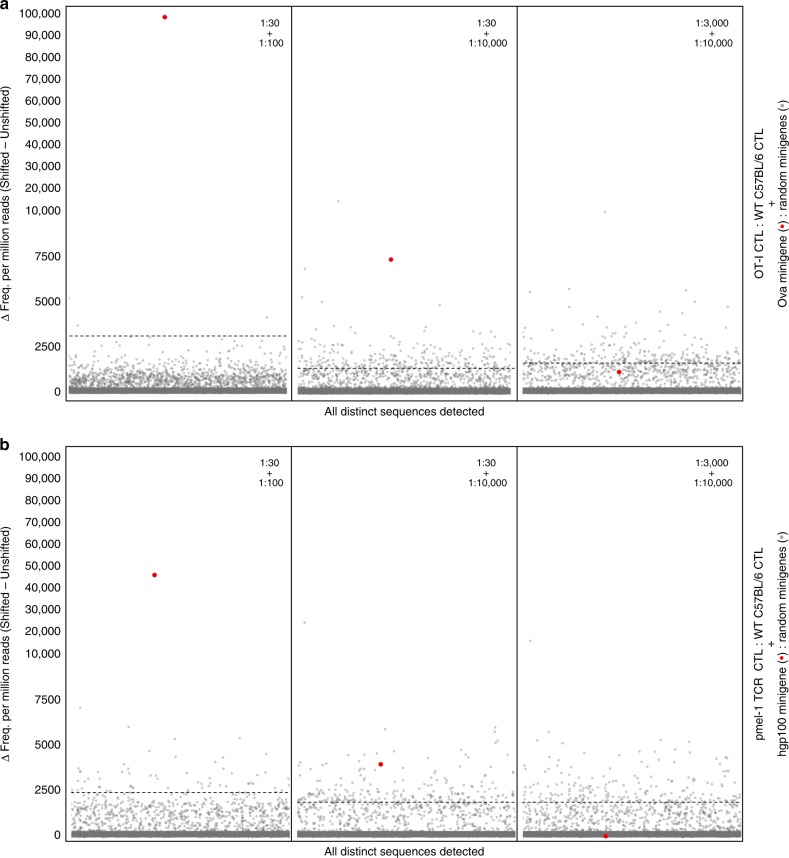
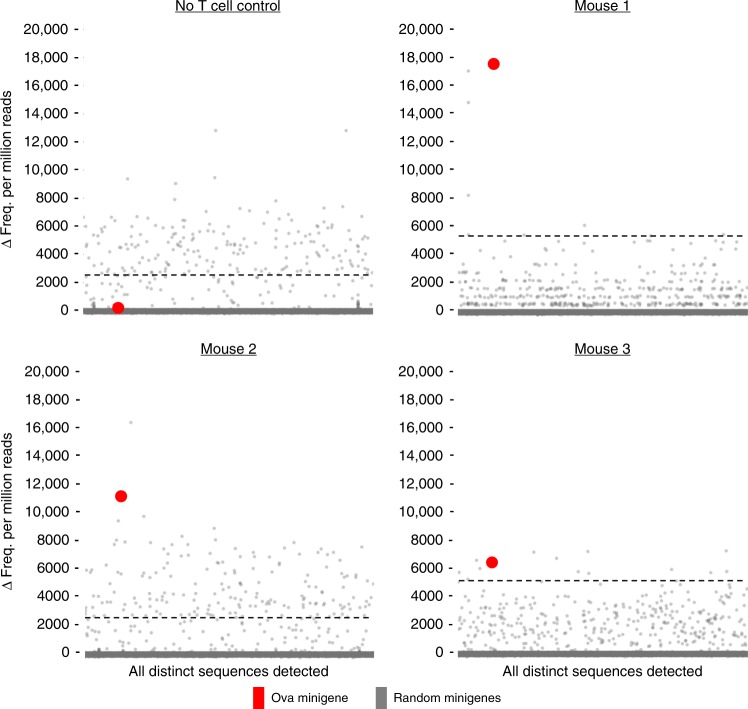
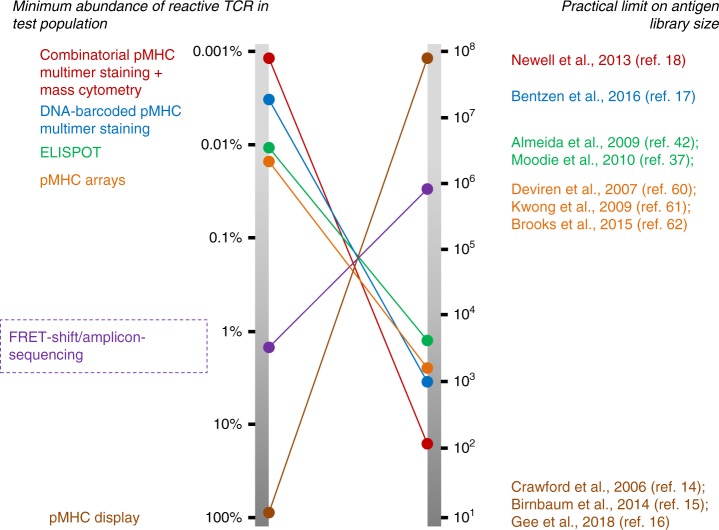
Cytotoxic CD8+ T cells recognize and eliminate infected or malignant cells that present peptide epitopes derived from intracellularly processed antigens on their surface. However, comprehensive profiling of specific major histocompatibility complex (MHC)-bound peptide epitopes that are naturally processed and capable of eliciting a functional T cell response has been challenging. Here, we report a method for deep and unbiased T cell epitope profiling, using in vitro co-culture of CD8+ T cells together with target cells transduced with high-complexity, epitope-encoding minigene libraries. Target cells that are subject to cytotoxic attack from T cells in co-culture are isolated prior to apoptosis by fluorescence-activated cell sorting, and characterized by sequencing the encoded minigenes. We then validate this highly parallelized method using known murine T cell receptor/peptide-MHC pairs and diverse minigene-encoded epitope libraries. Our data thus suggest that this epitope profiling method allows unambiguous and sensitive identification of naturally processed and MHC-presented peptide epitopes.
Conflict of interest statement
G.S. and R.A.H. are named inventors on patent applications, filed by the British Columbia Cancer Agency, encompassing the methods presented here: PCT/CA2015/050230, CA2943569A, and US20180052176A1 (priority date 2014-03-27).
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
