

A Genome-Wide Knockout Screen in Human Macrophages Identified Host Factors Modulating Salmonella Infection
- PMID: 31594818
- PMCID: PMC6786873
- DOI: 10.1128/mBio.02169-19
A Genome-Wide Knockout Screen in Human Macrophages Identified Host Factors Modulating Salmonella Infection
Abstract
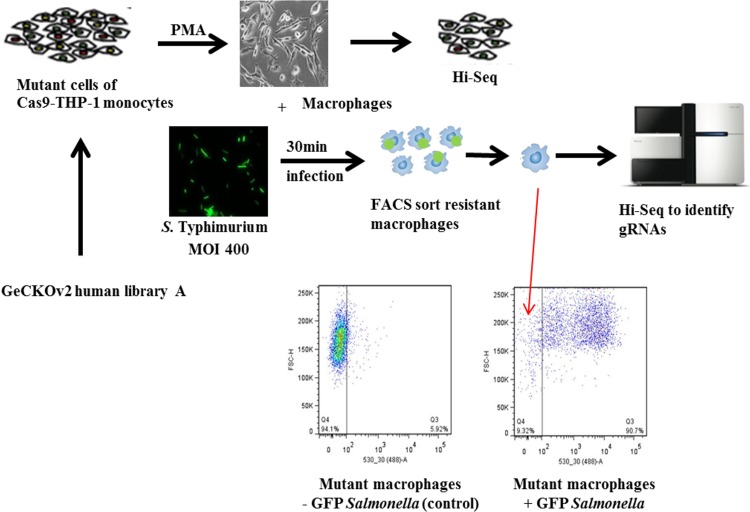
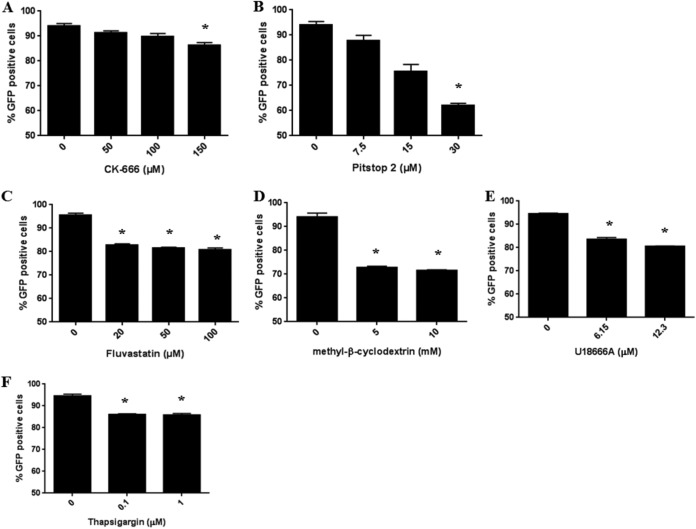
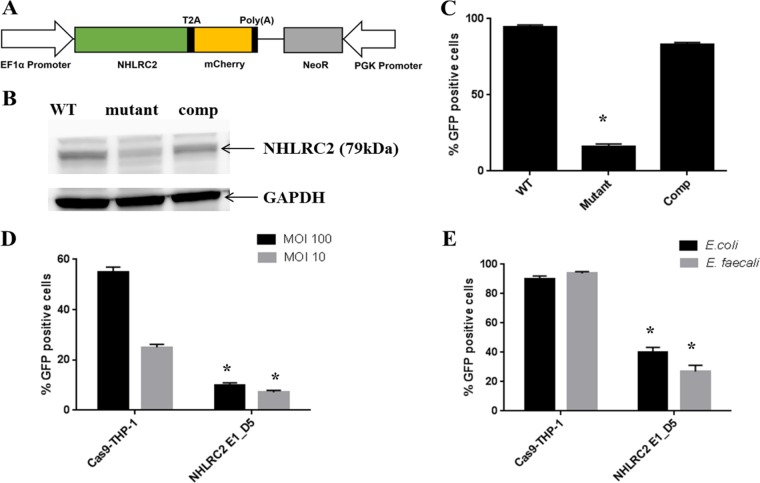
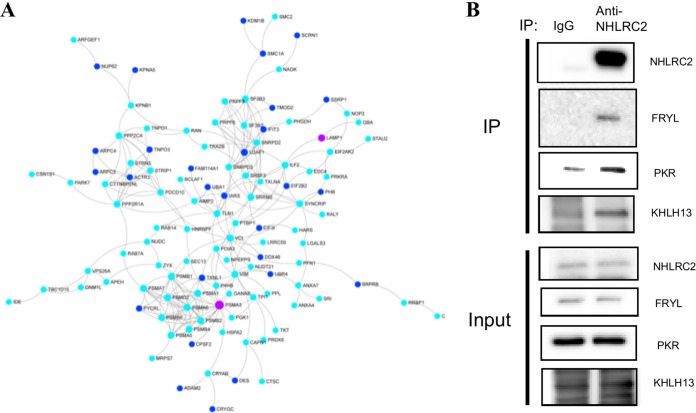
A genome-scale CRISPR knockout library screen of THP-1 human macrophages was performed to identify loss-of-function mutations conferring resistance to Salmonella uptake. The screen identified 183 candidate genes, from which 14 representative genes involved in actin dynamics (ACTR3, ARPC4, CAPZB, TOR3A, CYFIP2, CTTN, and NHLRC2), glycosaminoglycan metabolism (B3GNT1), receptor signaling (PDGFB and CD27), lipid raft formation (CLTCL1), calcium transport (ATP2A2 and ITPR3), and cholesterol metabolism (HMGCR) were analyzed further. For some of these pathways, known chemical inhibitors could replicate the Salmonella resistance phenotype, indicating their potential as targets for host-directed therapy. The screen indicated a role for the relatively uncharacterized gene NHLRC2 in both Salmonella invasion and macrophage differentiation. Upon differentiation, NHLRC2 mutant macrophages were hyperinflammatory and did not exhibit characteristics typical of macrophages, including atypical morphology and inability to interact and phagocytose bacteria/particles. Immunoprecipitation confirmed an interaction of NHLRC2 with FRYL, EIF2AK2, and KLHL13.IMPORTANCESalmonella exploits macrophages to gain access to the lymphatic system and bloodstream to lead to local and potentially systemic infections. With an increasing number of antibiotic-resistant isolates identified in humans, Salmonella infections have become major threats to public health. Therefore, there is an urgent need to identify alternative approaches to anti-infective therapy, including host-directed therapies. In this study, we used a simple genome-wide screen to identify 183 candidate host factors in macrophages that can confer resistance to Salmonella infection. These factors may be potential therapeutic targets against Salmonella infections.
Keywords: CRISPR; Salmonella; bacteria; genome-wide screen; macrophages.
Copyright © 2019 Yeung et al.
Figures



References
-
- Eng S-K, Pusparajah P, Ab Mutalib N-S, Ser H-L, Chan K-G, Lee L-H. 2015. Salmonella: a review on pathogenesis, epidemiology and antibiotic resistance. Front Life Sci 8:284–293. doi: 10.1080/21553769.2015.1051243. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
