

Glucoselysine is derived from fructose and accumulates in the eye lens of diabetic rats
- PMID: 31594865
- PMCID: PMC6873178
- DOI: 10.1074/jbc.RA119.010744
Glucoselysine is derived from fructose and accumulates in the eye lens of diabetic rats
Abstract
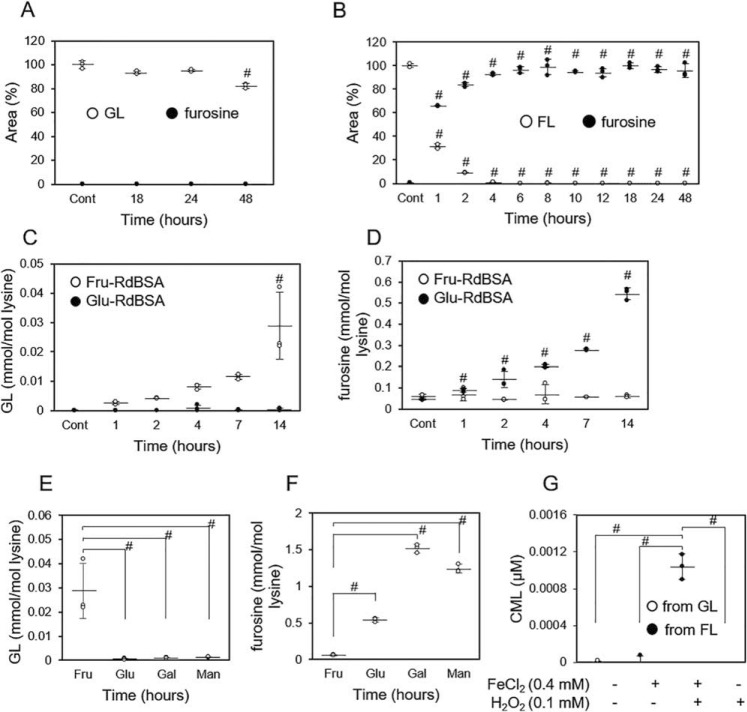
Prolonged hyperglycemia generates advanced glycation end-products (AGEs), which are believed to be involved in the pathogenesis of diabetic complications. In the present study, we developed a polyclonal antibody against fructose-modified proteins (Fru-P antibody) and identified its epitope as glucoselysine (GL) by NMR and LC-electrospray ionization (ESI)- quadrupole TOF (QTOF) analyses and evaluated its potential role in diabetes sequelae. Although the molecular weight of GL was identical to that of fructoselysine (FL), GL was distinguishable from FL because GL was resistant to acid hydrolysis, which converted all of the FLs to furosine. We also detected GL in vitro when reduced BSA was incubated with fructose for 1 day. However, when we incubated reduced BSA with glucose, galactose, or mannose for 14 days, we did not detect GL, suggesting that GL is dominantly generated from fructose. LC-ESI-MS/MS experiments with synthesized [13C6]GL indicated that the GL levels in the rat eye lens time-dependently increase after streptozotocin-induced diabetes. We observed a 31.3-fold increase in GL 8 weeks after the induction compared with nondiabetic rats, and Nϵ-(carboxymethyl)lysine and furosine increased by 1.7- and 21.5-fold, respectively, under the same condition. In contrast, sorbitol in the lens levelled off at 2 weeks after diabetes induction. We conclude that GL may be a useful biological marker to monitor and elucidate the mechanism of protein degeneration during progression of diabetes.
Keywords: AGEs; diabetes; glucoselysine; glycation; lens; mass spectrometry (MS); nuclear magnetic resonance (NMR); polyclonal antibody; polyol pathway; post-translational modification (PTM).
© 2019 Ohno et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article.
Figures






References
-
- Schleicher E. D., Gerbitz K. D., Dolhofer R., Reindl E., Wieland O. H., Edelmann E., Haslbeck M., Kemmler W., Walter H., and Mehnert H. (1984) Clinical utility of nonenzymatically glycosylated blood proteins as an index of glucose control. Diabetes Care 7, 548–556 10.2337/diacare.7.6.548 - DOI - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
