

A Perspective: Active Role of Lipids in Neurotransmitter Dynamics
- PMID: 31595461
- PMCID: PMC7031182
- DOI: 10.1007/s12035-019-01775-7
A Perspective: Active Role of Lipids in Neurotransmitter Dynamics
Abstract
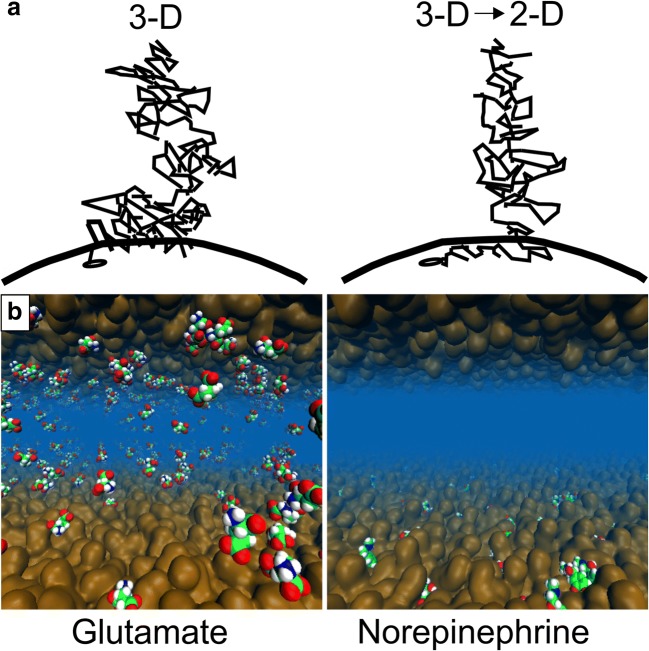
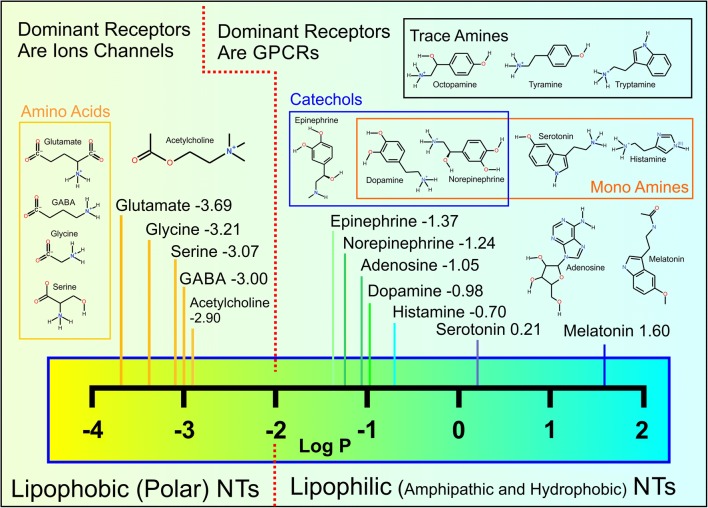
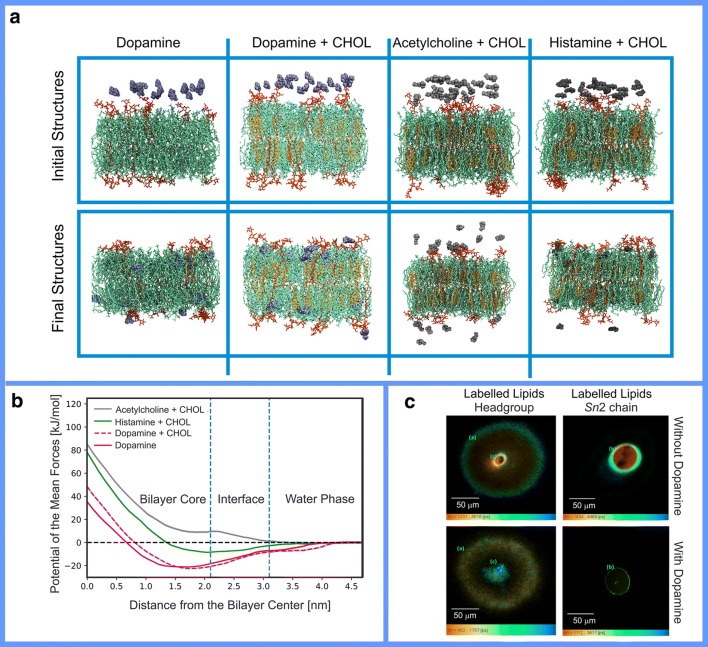
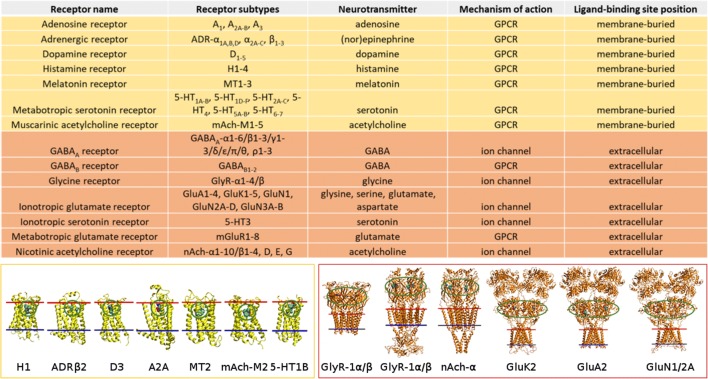
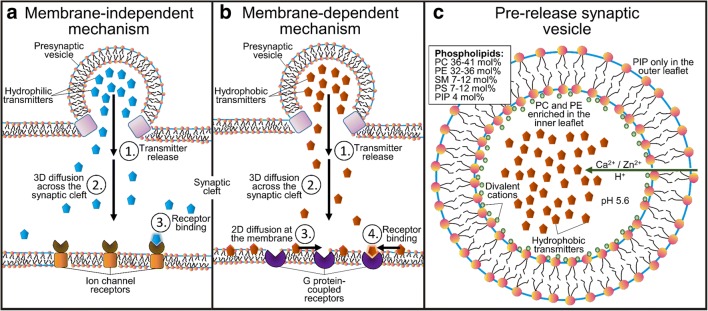
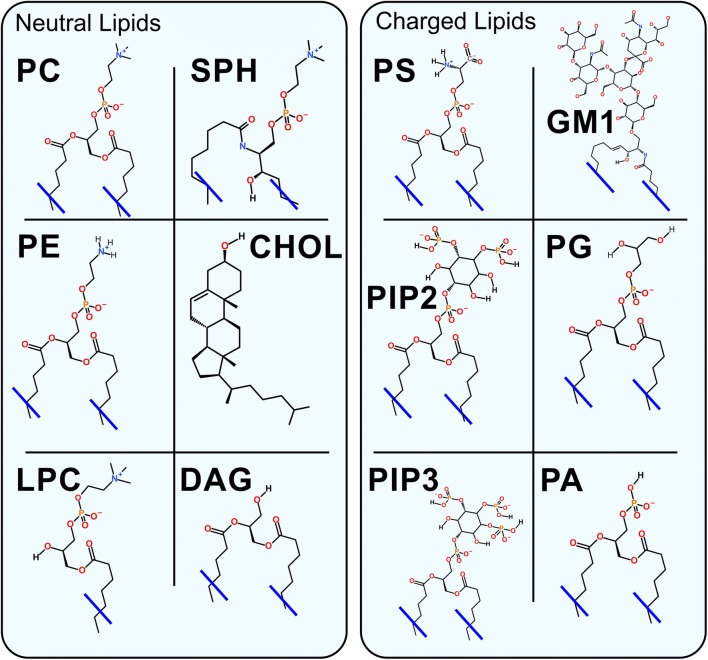
Synaptic neurotransmission is generally considered as a function of membrane-embedded receptors and ion channels in response to the neurotransmitter (NT) release and binding. This perspective aims to widen the protein-centric view by including another vital component-the synaptic membrane-in the discussion. A vast set of atomistic molecular dynamics simulations and biophysical experiments indicate that NTs are divided into membrane-binding and membrane-nonbinding categories. The binary choice takes place at the water-membrane interface and follows closely the positioning of the receptors' binding sites in relation to the membrane. Accordingly, when a lipophilic NT is on route to a membrane-buried binding site, it adheres on the membrane and, then, travels along its plane towards the receptor. In contrast, lipophobic NTs, which are destined to bind into receptors with extracellular binding sites, prefer the water phase. This membrane-based sorting splits the neurotransmission into membrane-independent and membrane-dependent mechanisms and should make the NT binding into the receptors more efficient than random diffusion would allow. The potential implications and notable exceptions to the mechanisms are discussed here. Importantly, maintaining specific membrane lipid compositions (MLCs) at the synapses, especially regarding anionic lipids, affect the level of NT-membrane association. These effects provide a plausible link between the MLC imbalances and neurological diseases such as depression or Parkinson's disease. Moreover, the membrane plays a vital role in other phases of the NT life cycle, including storage and release from the synaptic vesicles, transport from the synaptic cleft, as well as their synthesis and degradation.
Keywords: Membrane lipid composition (MLC); Membrane-based sorting; Molecular dynamics (MD); Neurotransmitter; Synaptic neurotransmission; Synaptic receptor.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Zucker RS, Kullmann DM, Kaeser PS (2014) Release of Neurotransmitters. In: Byrne JH, Heidelberger R, Waxham MN (eds) From Molecules to Networks. Elsevier Inc., pp 443–488
-
- Deutch AY, Roth RH (2014) Pharmacology and Biochemistry of Synaptic Transmission Classical Transmitters. In: Byrne JH, Heidelberger R, Waxham MN (eds) From Molecules to Networks. Elsevier Inc., pp 207–237
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
