

Bisphenol AF promotes inflammation in human white adipocytes
- PMID: 31596606
- PMCID: PMC6985838
- DOI: 10.1152/ajpcell.00175.2019
Bisphenol AF promotes inflammation in human white adipocytes
Abstract
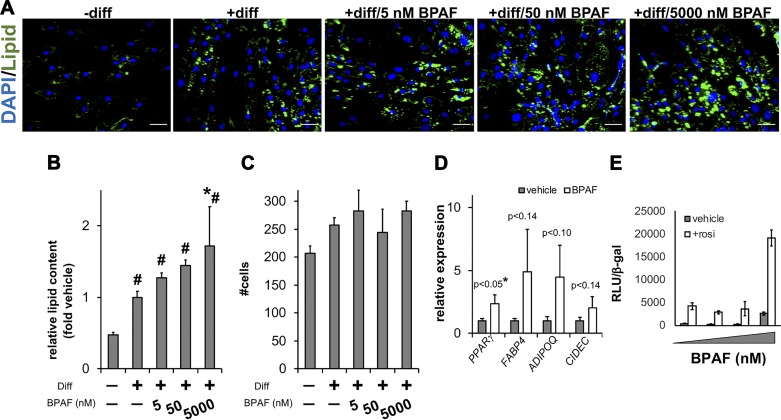
Endocrine-disrupting chemicals interact with transcription factors essential for adipocyte differentiation. Exposure to endocrine-disrupting chemicals corresponds with elevated risks of obesity, but the effects of these compounds on human cells remain largely undefined. Widespread use of bisphenol AF (BPAF) as a bisphenol A (BPA) alternative in the plastics industry presents unknown health risks. To this end, we discovered that BPAF interferes with the metabolic function of mature human adipocytes. Although 4-day exposures to BPAF accelerated adipocyte differentiation, we observed no effect on mature fat cell marker genes. Additional gene and protein expression analysis showed that BPAF treatment during human adipocyte differentiation failed to suppress the proinflammatory transcription factor STAT1. Microscopy and respirometry experiments demonstrated that BPAF impaired mitochondrial function and structure. To test the hypothesis that BPAF fosters vulnerabilities to STAT1 activation, we treated mature adipocytes previously exposed to BPAF with interferon-γ (IFNγ). BPAF increased IFNγ activation of STAT1 and exposed mitochondrial vulnerabilities that disrupt adipocyte lipid and carbohydrate metabolism. Collectively, our data establish that BPAF activates inflammatory signaling pathways that degrade metabolic activity in human adipocytes. These findings suggest how the BPA alternative BPAF contributes to metabolic changes that correspond with obesity.
Keywords: PPARγ; adipocyte; inflammation; metabolism; transcription.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures




References
-
- Adams M, Montague CT, Prins JB, Holder JC, Smith SA, Sanders L, Digby JE, Sewter CP, Lazar MA, Chatterjee VK, O’Rahilly S. Activators of peroxisome proliferator-activated receptor gamma have depot-specific effects on human preadipocyte differentiation. J Clin Invest 100: 3149–3153, 1997. doi:10.1172/JCI119870. - DOI - PMC - PubMed
-
- Arancio AL, Cole KD, Dominguez AR, Cohenour ER, Kadie J, Maloney WC, Cilliers C, Schuh SM. Bisphenol A, Bisphenol AF, di-n-butyl phthalate, and 17β-estradiol have shared and unique dose-dependent effects on early embryo cleavage divisions and development in Xenopus laevis. Reprod Toxicol 84: 65–74, 2019. doi:10.1016/j.reprotox.2018.12.005. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous