

Phosphoregulation of tropomyosin is crucial for actin cable turnover and division site placement
- PMID: 31597679
- PMCID: PMC6829654
- DOI: 10.1083/jcb.201809089
Phosphoregulation of tropomyosin is crucial for actin cable turnover and division site placement
Abstract
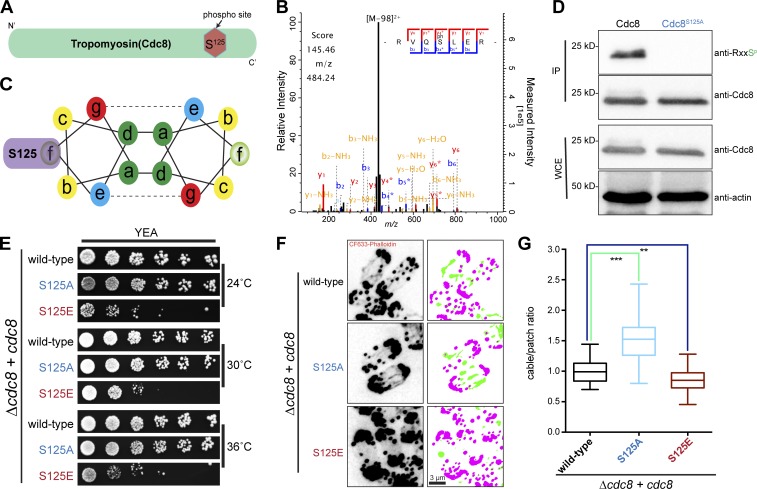
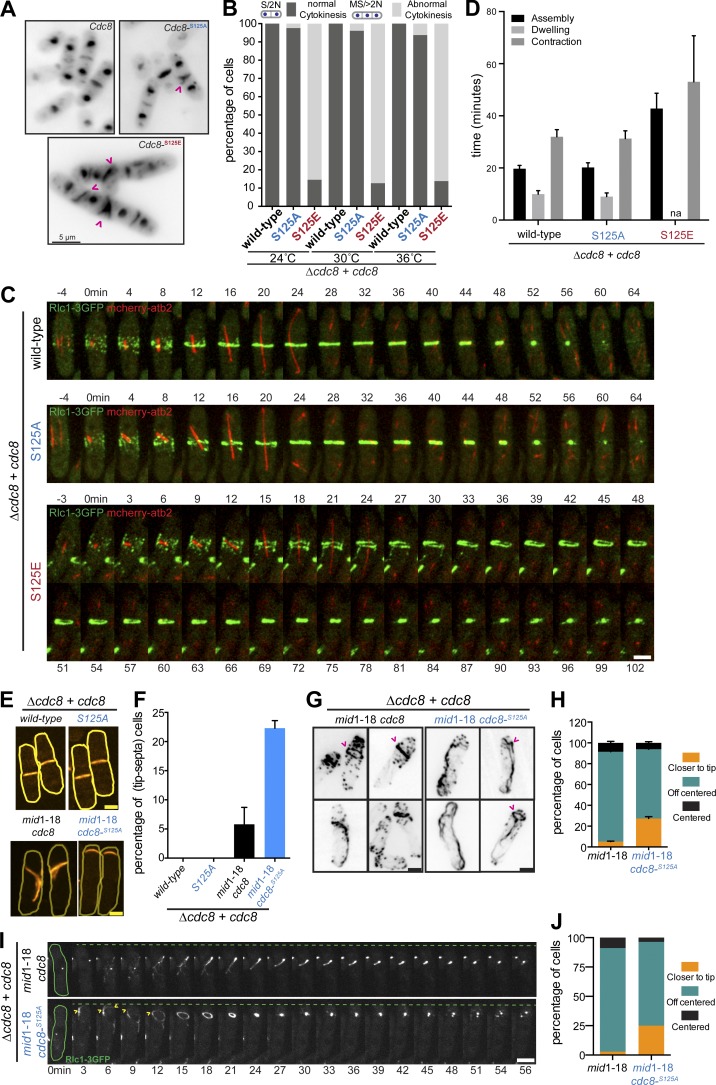
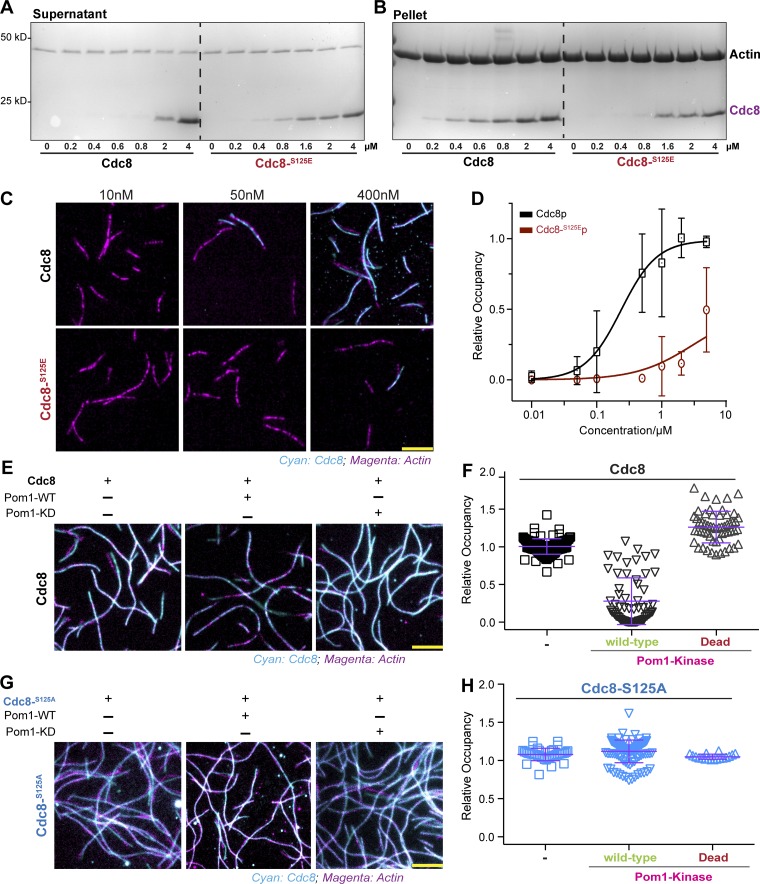
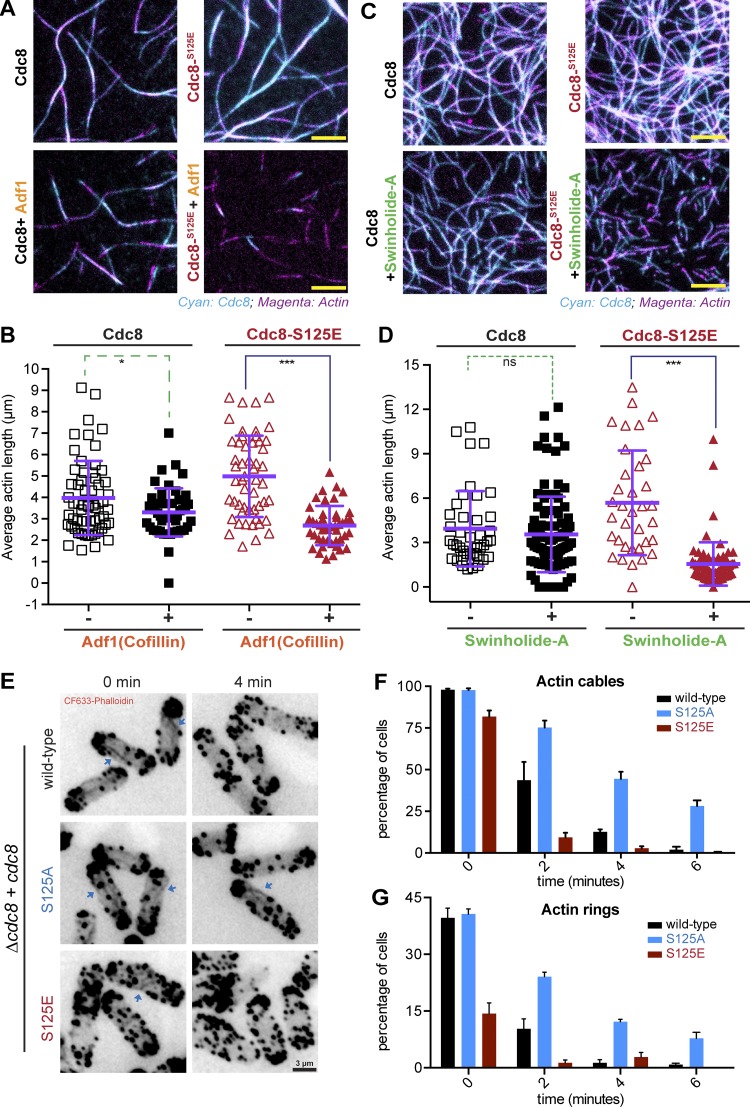
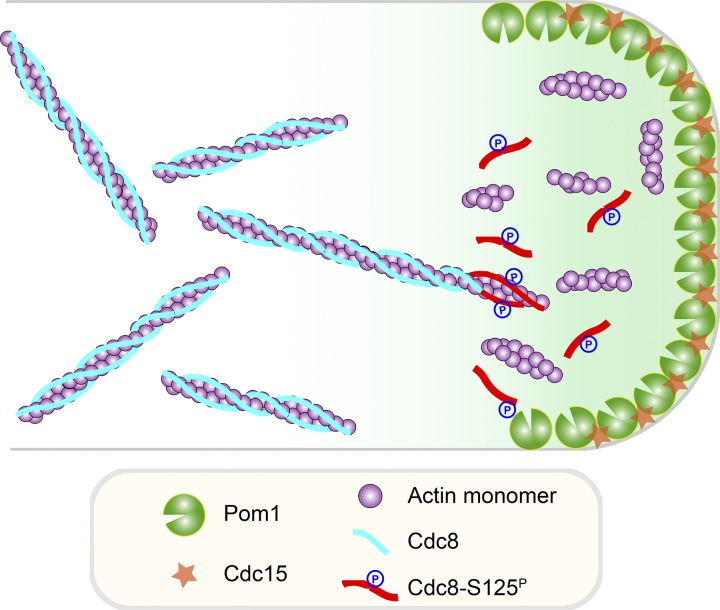
Tropomyosin is a coiled-coil actin binding protein key to the stability of actin filaments. In muscle cells, tropomyosin is subject to calcium regulation, but its regulation in nonmuscle cells is not understood. Here, we provide evidence that the fission yeast tropomyosin, Cdc8, is regulated by phosphorylation of a serine residue. Failure of phosphorylation leads to an increased number and stability of actin cables and causes misplacement of the division site in certain genetic backgrounds. Phosphorylation of Cdc8 weakens its interaction with actin filaments. Furthermore, we show through in vitro reconstitution that phosphorylation-mediated release of Cdc8 from actin filaments facilitates access of the actin-severing protein Adf1 and subsequent filament disassembly. These studies establish that phosphorylation may be a key mode of regulation of nonmuscle tropomyosins, which in fission yeast controls actin filament stability and division site placement.
© 2019 Palani et al.
Figures
Similar articles
-
Acetylation regulates tropomyosin function in the fission yeast Schizosaccharomyces pombe.J Cell Sci. 2007 May 1;120(Pt 9):1635-45. doi: 10.1242/jcs.001115. J Cell Sci. 2007. PMID: 17452625
-
Phosphoregulation of tropomyosin-actin interaction revealed using a genetic code expansion strategy.Wellcome Open Res. 2020 Jul 7;5:161. doi: 10.12688/wellcomeopenres.16082.1. eCollection 2020. Wellcome Open Res. 2020. PMID: 32802966 Free PMC article.
-
Altering the stability of the Cdc8 overlap region modulates the ability of this tropomyosin to bind co-operatively to actin and regulate myosin.Biochem J. 2011 Sep 1;438(2):265-73. doi: 10.1042/BJ20101316. Biochem J. 2011. PMID: 21658004
-
Periodicities designed in the tropomyosin sequence and structure define its functions.Bioarchitecture. 2013 May-Jun;3(3):51-6. doi: 10.4161/bioa.25616. Epub 2013 Jul 8. Bioarchitecture. 2013. PMID: 23887197 Free PMC article. Review.
-
New insights into the regulation of the actin cytoskeleton by tropomyosin.Int Rev Cell Mol Biol. 2010;281:91-128. doi: 10.1016/S1937-6448(10)81003-2. Int Rev Cell Mol Biol. 2010. PMID: 20460184 Free PMC article. Review.
Cited by
-
Novel human cell expression method reveals the role and prevalence of posttranslational modification in nonmuscle tropomyosins.J Biol Chem. 2021 Oct;297(4):101154. doi: 10.1016/j.jbc.2021.101154. Epub 2021 Sep 1. J Biol Chem. 2021. PMID: 34478714 Free PMC article.
-
Dynamics of Tpm1.8 domains on actin filaments with single-molecule resolution.Mol Biol Cell. 2020 Oct 15;31(22):2452-2462. doi: 10.1091/mbc.E19-10-0586. Epub 2020 Aug 26. Mol Biol Cell. 2020. PMID: 32845787 Free PMC article.
-
Acetylation of fission yeast tropomyosin does not promote differential association with cognate formins.Cytoskeleton (Hoboken). 2023 Mar;80(3-4):77-92. doi: 10.1002/cm.21745. Epub 2023 Feb 8. Cytoskeleton (Hoboken). 2023. PMID: 36692369 Free PMC article.
-
Visualizing the in vitro assembly of tropomyosin/actin filaments using TIRF microscopy.Biophys Rev. 2020 Aug;12(4):879-885. doi: 10.1007/s12551-020-00720-6. Epub 2020 Jul 7. Biophys Rev. 2020. PMID: 32638329 Free PMC article. Review.
-
Nfkb2 deficiency and its impact on plasma cells and immunoglobulin expression in murine small intestinal mucosa.Am J Physiol Gastrointest Liver Physiol. 2022 Oct 1;323(4):G306-G317. doi: 10.1152/ajpgi.00037.2022. Epub 2022 Aug 2. Am J Physiol Gastrointest Liver Physiol. 2022. PMID: 35916405 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
