

Migratory DCs activate TGF-β to precondition naïve CD8+ T cells for tissue-resident memory fate
- PMID: 31601741
- PMCID: PMC6939608
- DOI: 10.1126/science.aav5728
Migratory DCs activate TGF-β to precondition naïve CD8+ T cells for tissue-resident memory fate
Abstract
Epithelial resident memory T (eTRM) cells serve as sentinels in barrier tissues to guard against previously encountered pathogens. How eTRM cells are generated has important implications for efforts to elicit their formation through vaccination or prevent it in autoimmune disease. Here, we show that during immune homeostasis, the cytokine transforming growth factor β (TGF-β) epigenetically conditions resting naïve CD8+ T cells and prepares them for the formation of eTRM cells in a mouse model of skin vaccination. Naïve T cell conditioning occurs in lymph nodes (LNs), but not in the spleen, through major histocompatibility complex class I-dependent interactions with peripheral tissue-derived migratory dendritic cells (DCs) and depends on DC expression of TGF-β-activating αV integrins. Thus, the preimmune T cell repertoire is actively conditioned for a specialized memory differentiation fate through signals restricted to LNs.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Competing Interests:
The authors declare no competing interests
Figures
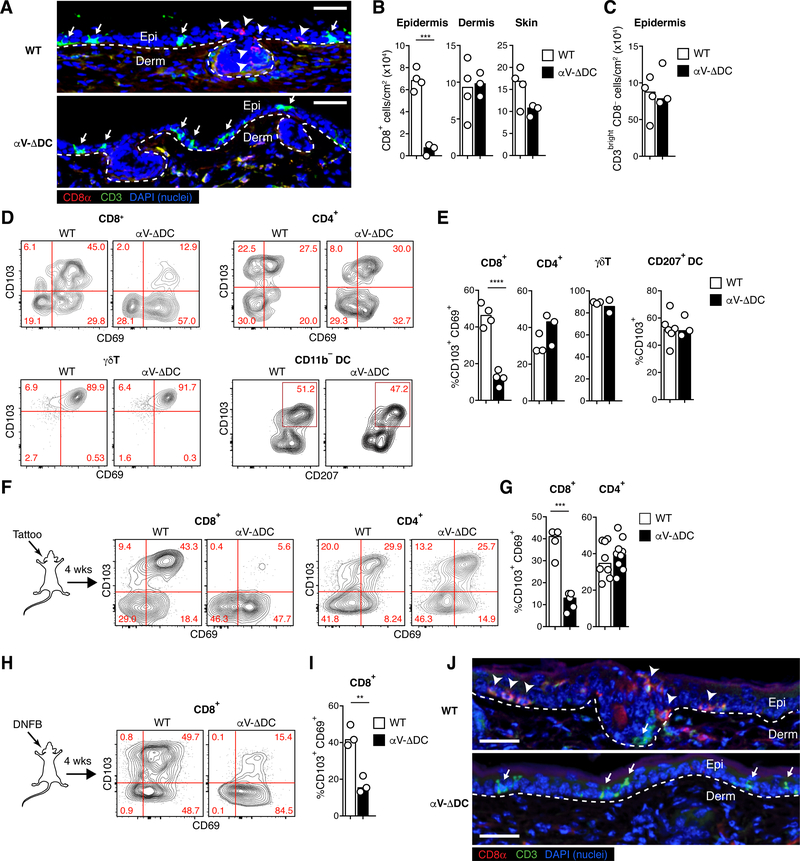
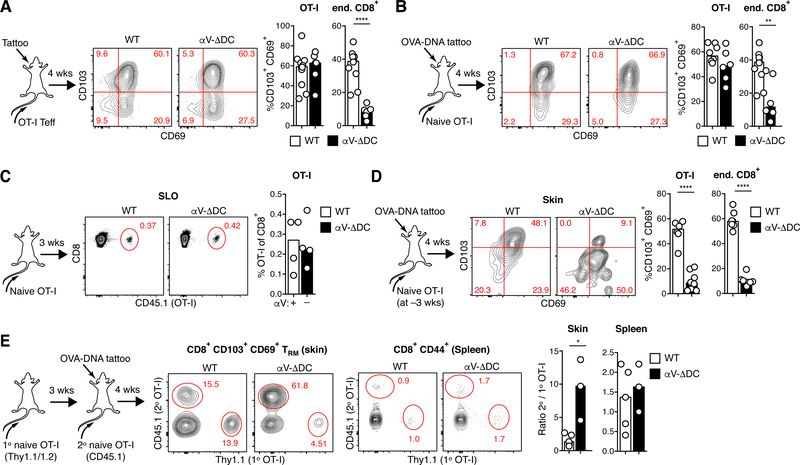
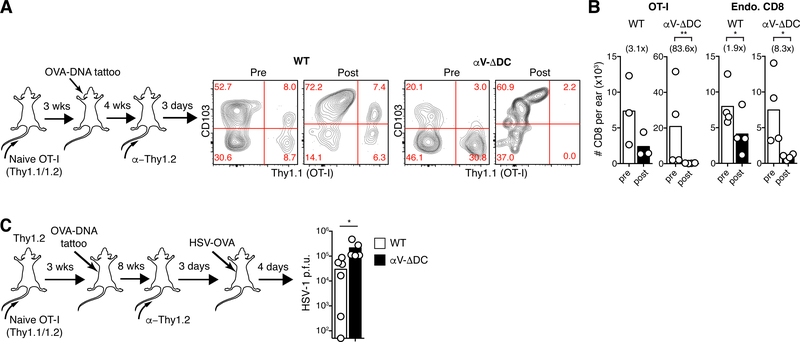
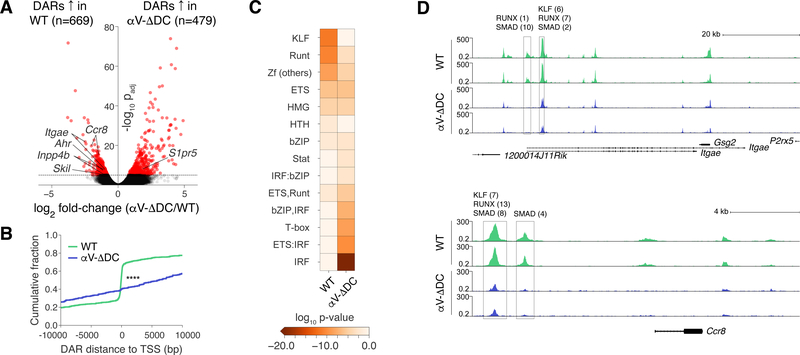
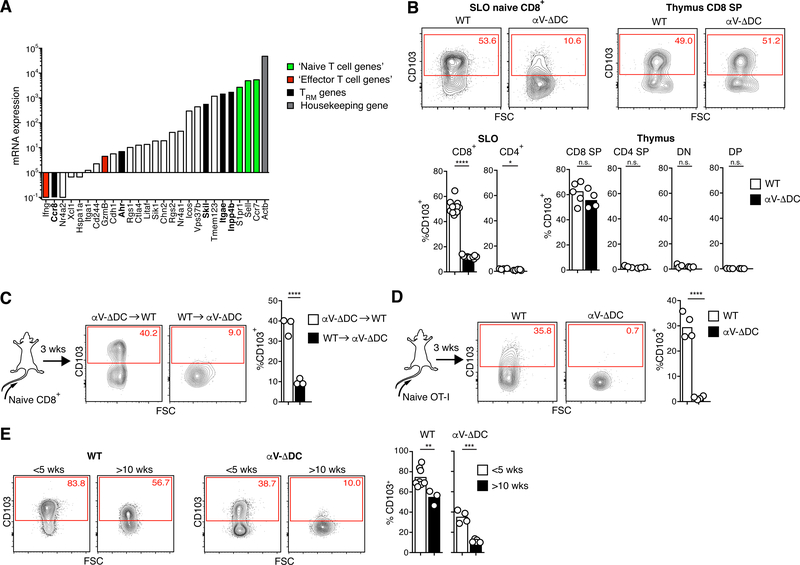
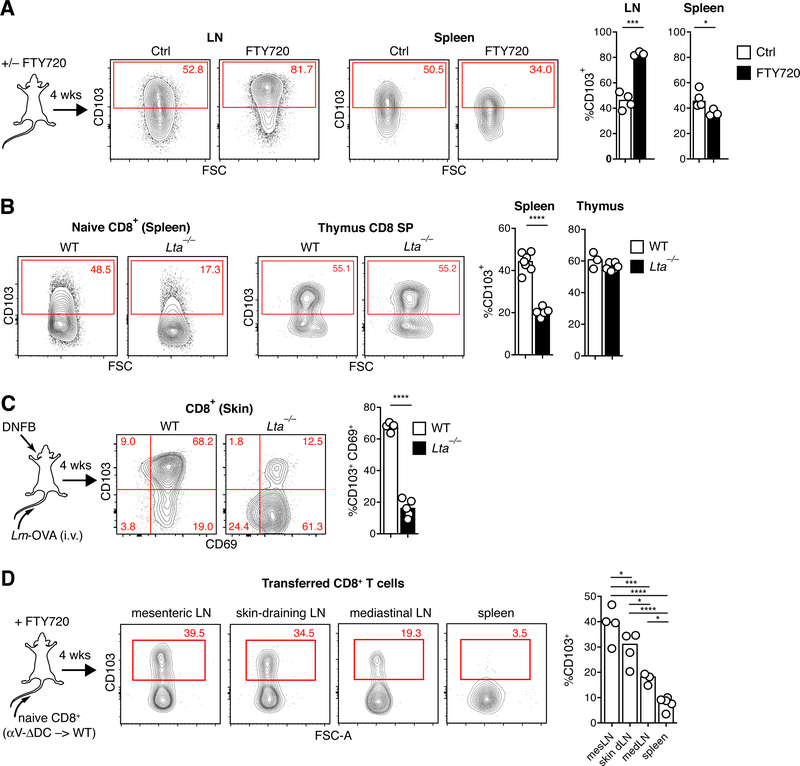
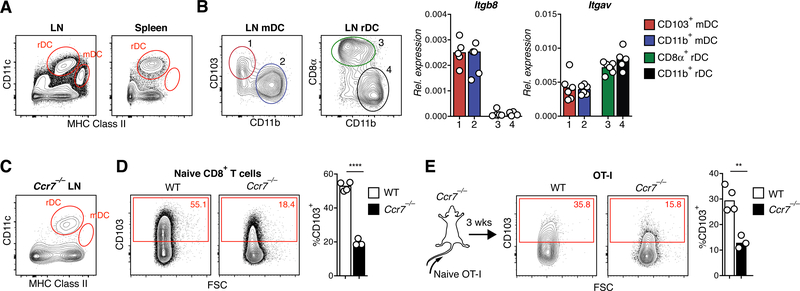
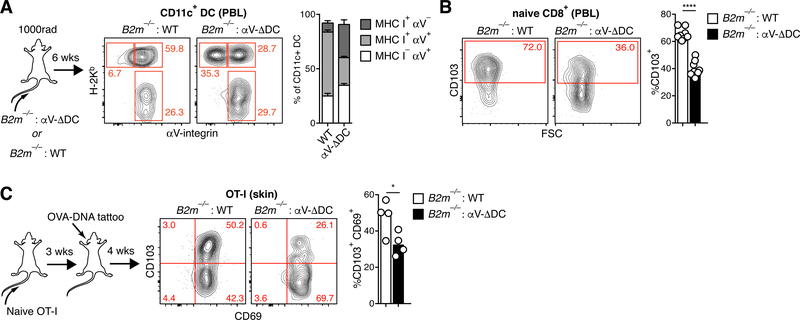
Comment in
-
Training T cells for tissue residence.Science. 2019 Oct 11;366(6462):188-189. doi: 10.1126/science.aaz3289. Science. 2019. PMID: 31601759 No abstract available.
-
Naive T cells can be primed for tissue residency.Nat Rev Immunol. 2019 Dec;19(12):718-719. doi: 10.1038/s41577-019-0239-z. Nat Rev Immunol. 2019. PMID: 31649326 No abstract available.
References
-
- Mueller SN, Mackay LK, Tissue-resident memory T cells: Local specialists in immune defence. Nat. Rev. Immunol. 16, 79–89 (2016). - PubMed
-
- Gebhardt T, Palendira U, Tscharke DC, Bedoui S, Tissue-resident memory T cells in tissue homeostasis, persistent infection, and cancer surveillance. Immunol. Rev. 283, 54–76 (2018). - PubMed
-
- Gebhardt T et al., Memory T cells in nonlymphoid tissue that provide enhanced local immunity during infection with herpes simplex virus. Nat. Immunol. 10, 524–530 (2009). - PubMed
-
- Schumacher TN et al., Skin-resident memory CD8+ T cells trigger a state of tissue-wide pathogen alert. Science (80-. ). 346, 101–105 (2014). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
