

Architecture of human Rag GTPase heterodimers and their complex with mTORC1
- PMID: 31601764
- PMCID: PMC6795536
- DOI: 10.1126/science.aax3939
Architecture of human Rag GTPase heterodimers and their complex with mTORC1
Abstract
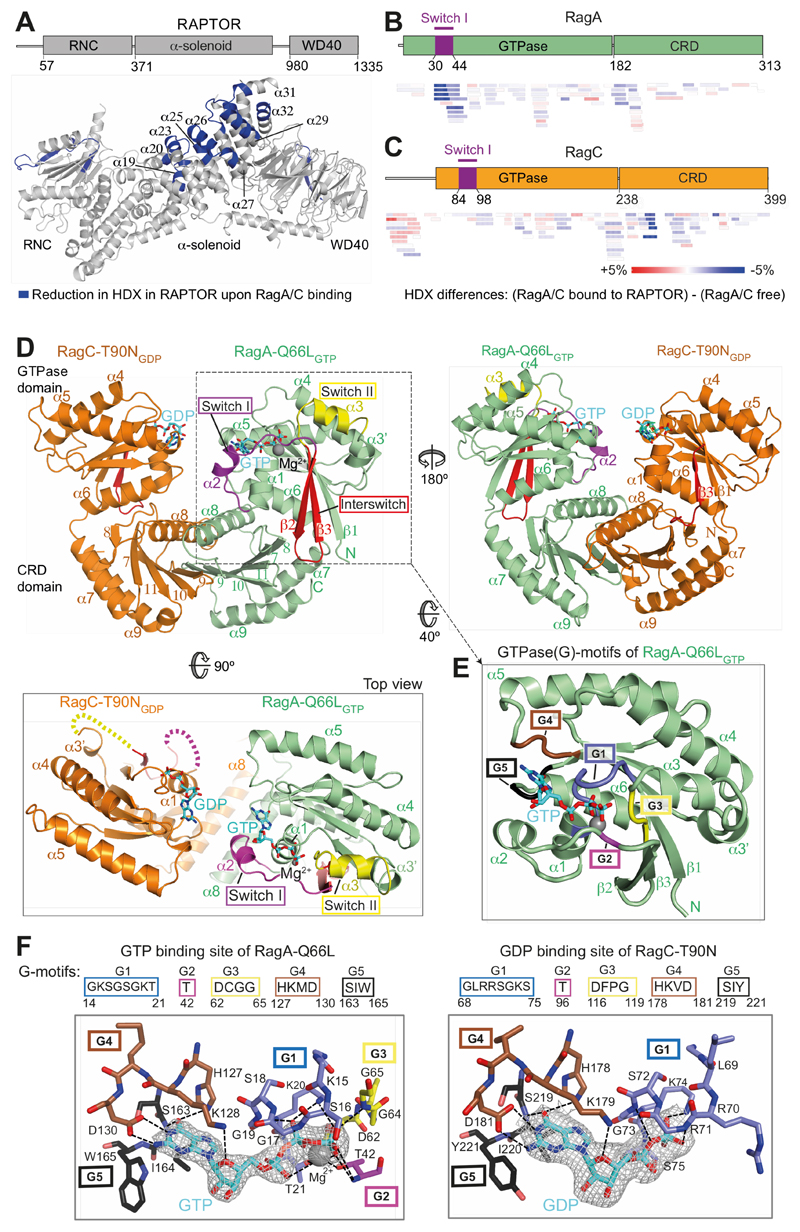
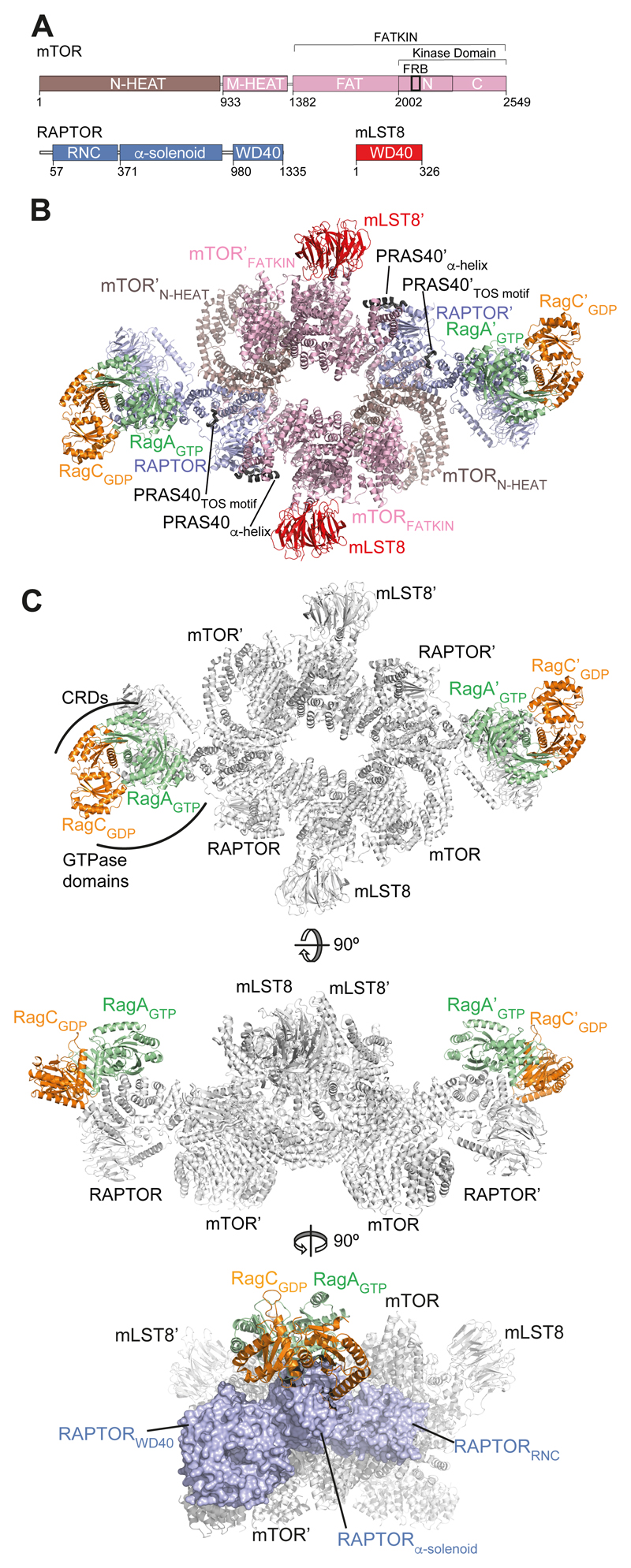
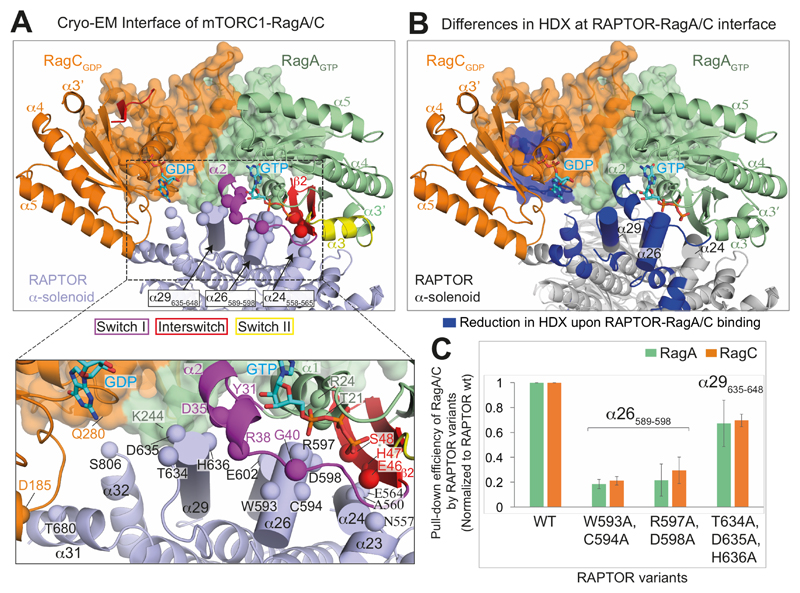
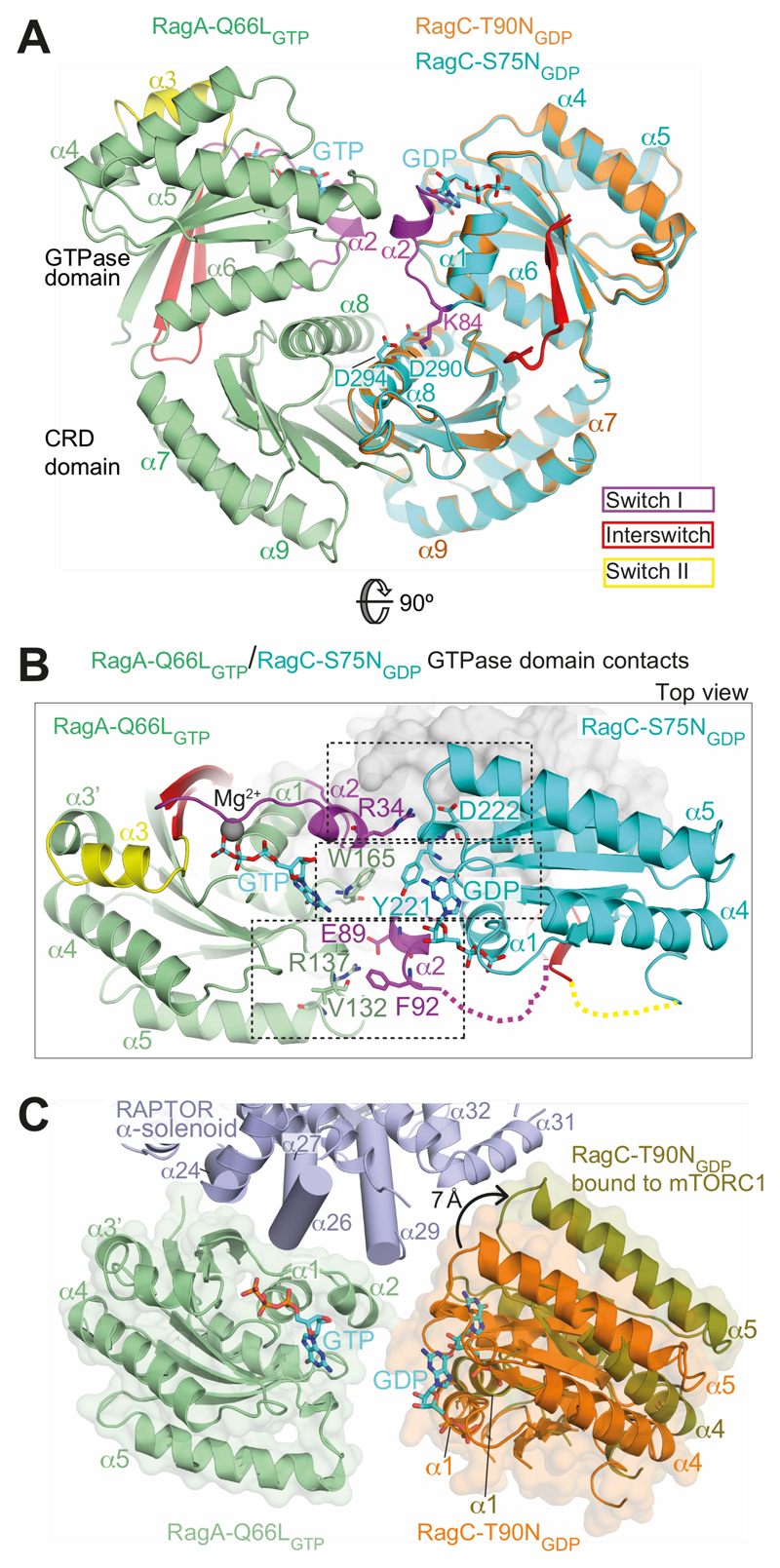
The Rag guanosine triphosphatases (GTPases) recruit the master kinase mTORC1 to lysosomes to regulate cell growth and proliferation in response to amino acid availability. The nucleotide state of Rag heterodimers is critical for their association with mTORC1. Our cryo-electron microscopy structure of RagA/RagC in complex with mTORC1 shows the details of RagA/RagC binding to the RAPTOR subunit of mTORC1 and explains why only the RagAGTP/RagCGDP nucleotide state binds mTORC1. Previous kinetic studies suggested that GTP binding to one Rag locks the heterodimer to prevent GTP binding to the other. Our crystal structures and dynamics of RagA/RagC show the mechanism for this locking and explain how oncogenic hotspot mutations disrupt this process. In contrast to allosteric activation by RHEB, Rag heterodimer binding does not change mTORC1 conformation and activates mTORC1 by targeting it to lysosomes.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
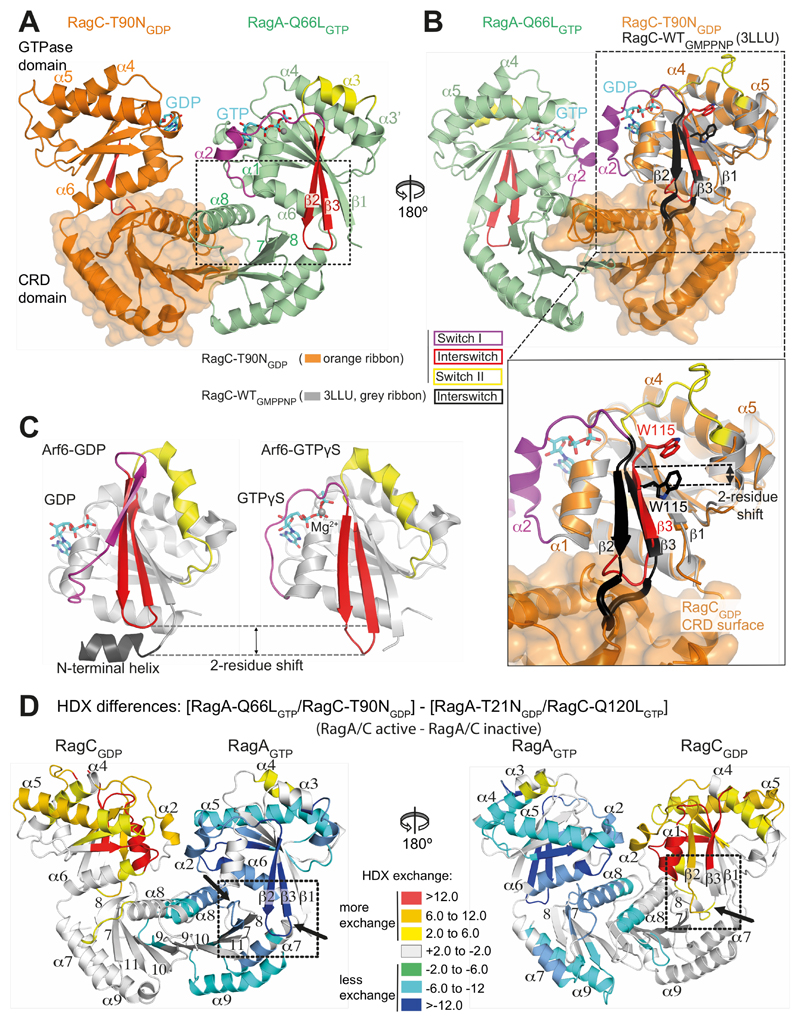
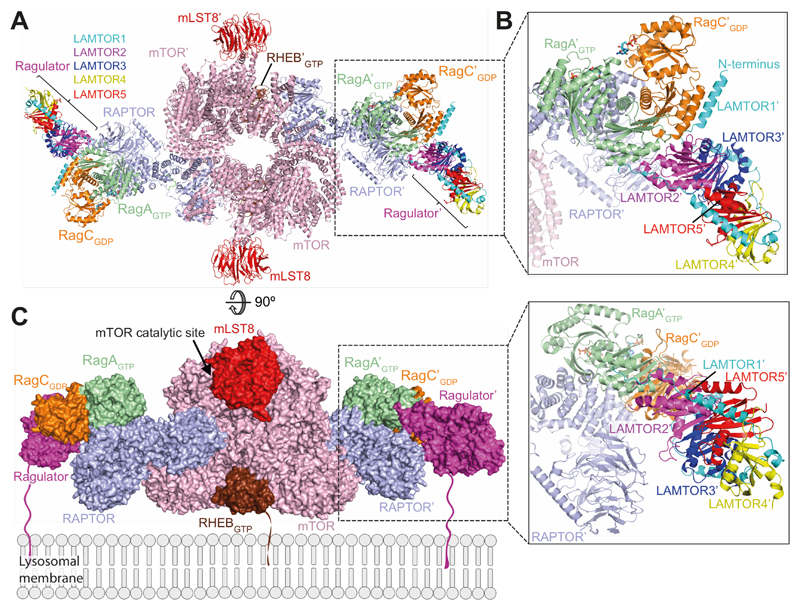
Comment in
-
Amino Acid Sensing: Architecture of mTORC1 on the Lysosome Surface.Curr Biol. 2020 Jan 20;30(2):R89-R91. doi: 10.1016/j.cub.2019.11.087. Curr Biol. 2020. PMID: 31962086
-
How mTORC1 makes sense of nutrients.Kidney Int. 2021 Feb;99(2):295-298. doi: 10.1016/j.kint.2020.07.052. Epub 2020 Oct 1. Kidney Int. 2021. PMID: 33010258 No abstract available.
References
-
- Cornu M, Albert V, Hall MN. mTOR in aging, metabolism, and cancer. Curr Opin Genet Dev. 2013;23:53–62. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
