

Keratin 19 regulates cell cycle pathway and sensitivity of breast cancer cells to CDK inhibitors
- PMID: 31601969
- PMCID: PMC6787034
- DOI: 10.1038/s41598-019-51195-9
Keratin 19 regulates cell cycle pathway and sensitivity of breast cancer cells to CDK inhibitors
Abstract
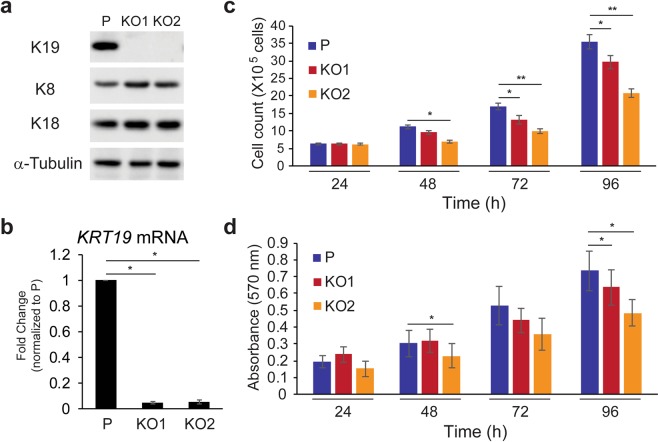
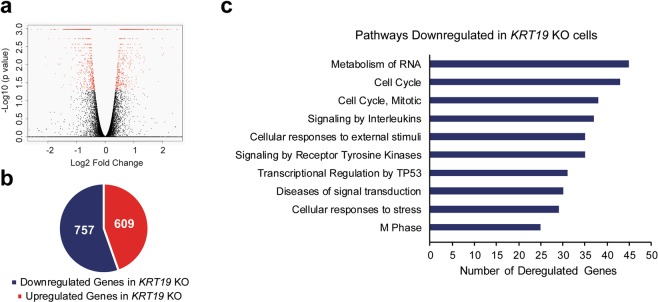
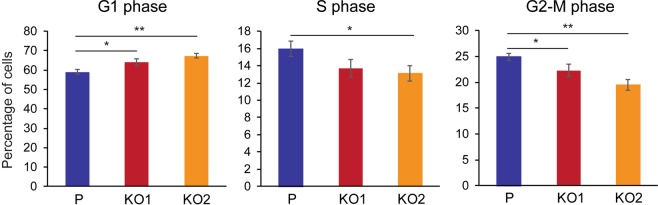
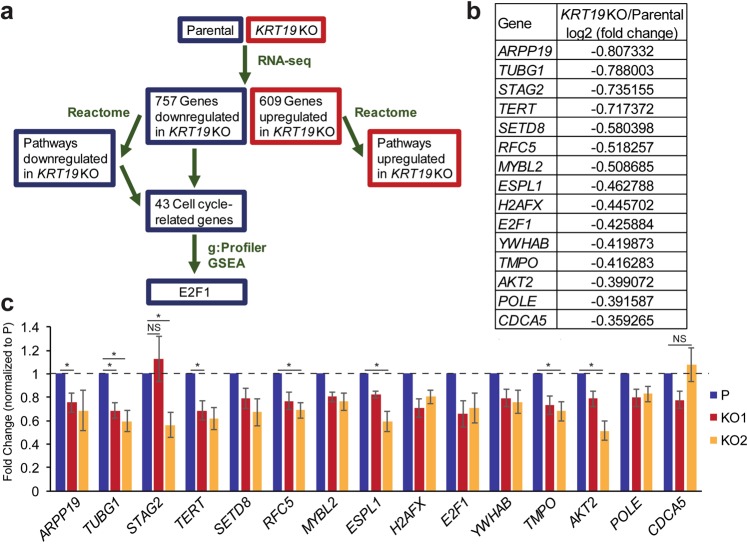
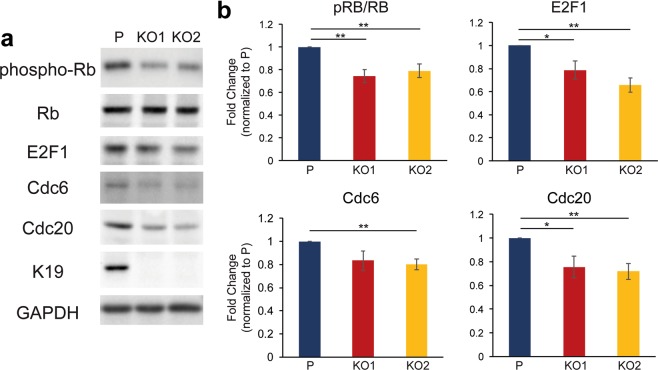
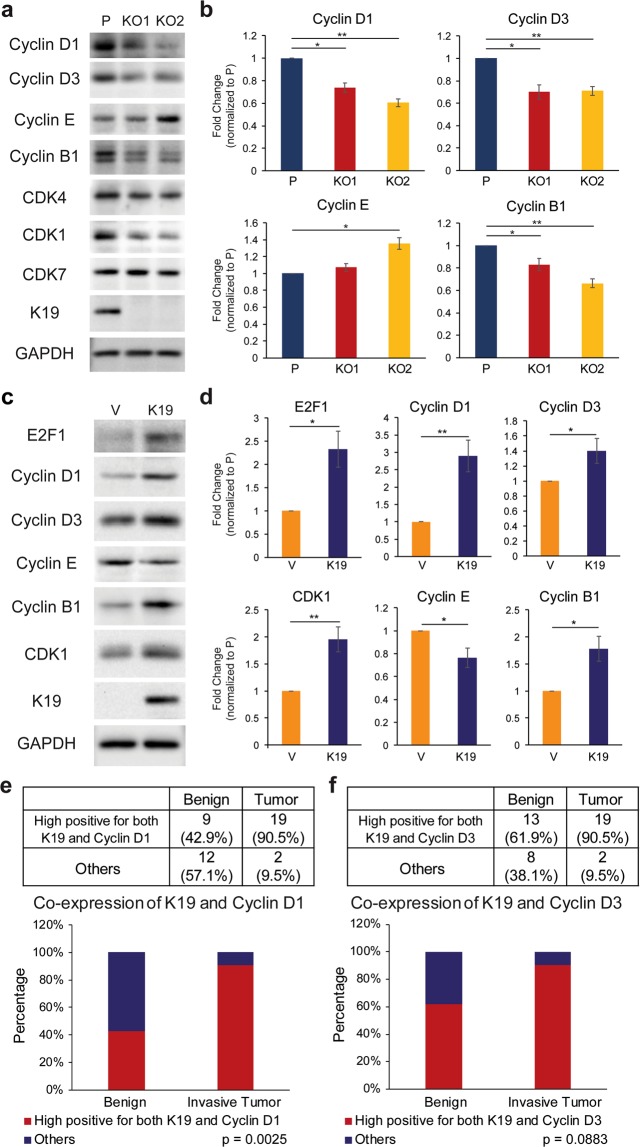
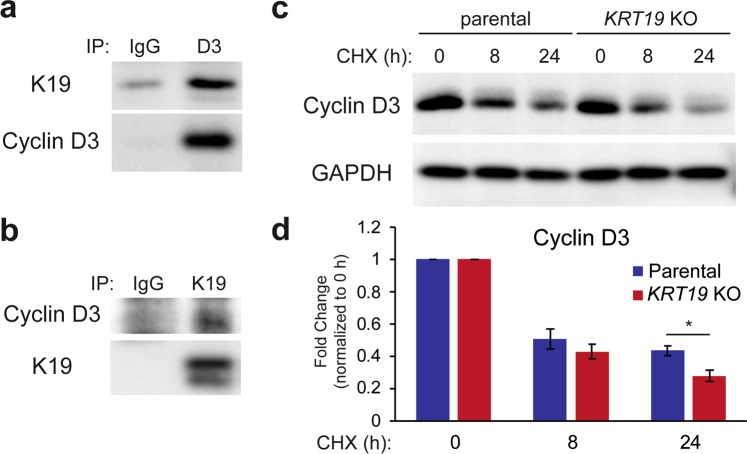
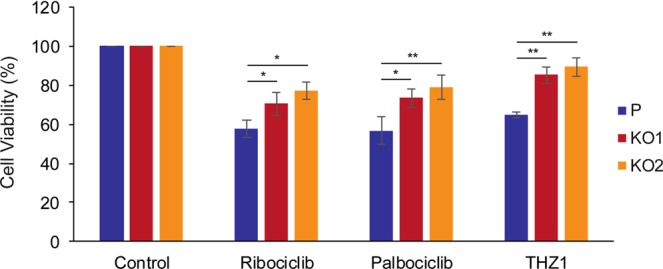
Keratin 19 (K19) belongs to the keratin family of proteins, which maintains structural integrity of epithelia. In cancer, K19 is highly expressed in several types where it serves as a diagnostic marker. Despite the positive correlation between higher expression of K19 in tumor and worse patient survival, the role of K19 in breast cancer remains unclear. Therefore, we ablated K19 expression in MCF7 breast cancer cells and found that K19 was required for cell proliferation. Transcriptome analyses of KRT19 knockout cells identified defects in cell cycle progression and levels of target genes of E2F1, a key transcriptional factor for the transition into S phase. Furthermore, proper levels of cyclin dependent kinases (CDKs) and cyclins, including D-type cyclins critical for E2F1 activation, were dependent on K19 expression, and K19-cyclin D co-expression was observed in human breast cancer tissues. Importantly, K19 interacts with cyclin D3, and a loss of K19 resulted in decreased protein stability of cyclin D3 and sensitivity of cells towards CDK inhibitor-induced cell death. Overall, these findings reveal a novel function of K19 in the regulation of cell cycle program and suggest that K19 may be used to predict the efficacy of CDK inhibitors for treatments of breast cancer.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
