

The intestinal microbiota programs diurnal rhythms in host metabolism through histone deacetylase 3
- PMID: 31604271
- PMCID: PMC7158748
- DOI: 10.1126/science.aaw3134
The intestinal microbiota programs diurnal rhythms in host metabolism through histone deacetylase 3
Abstract
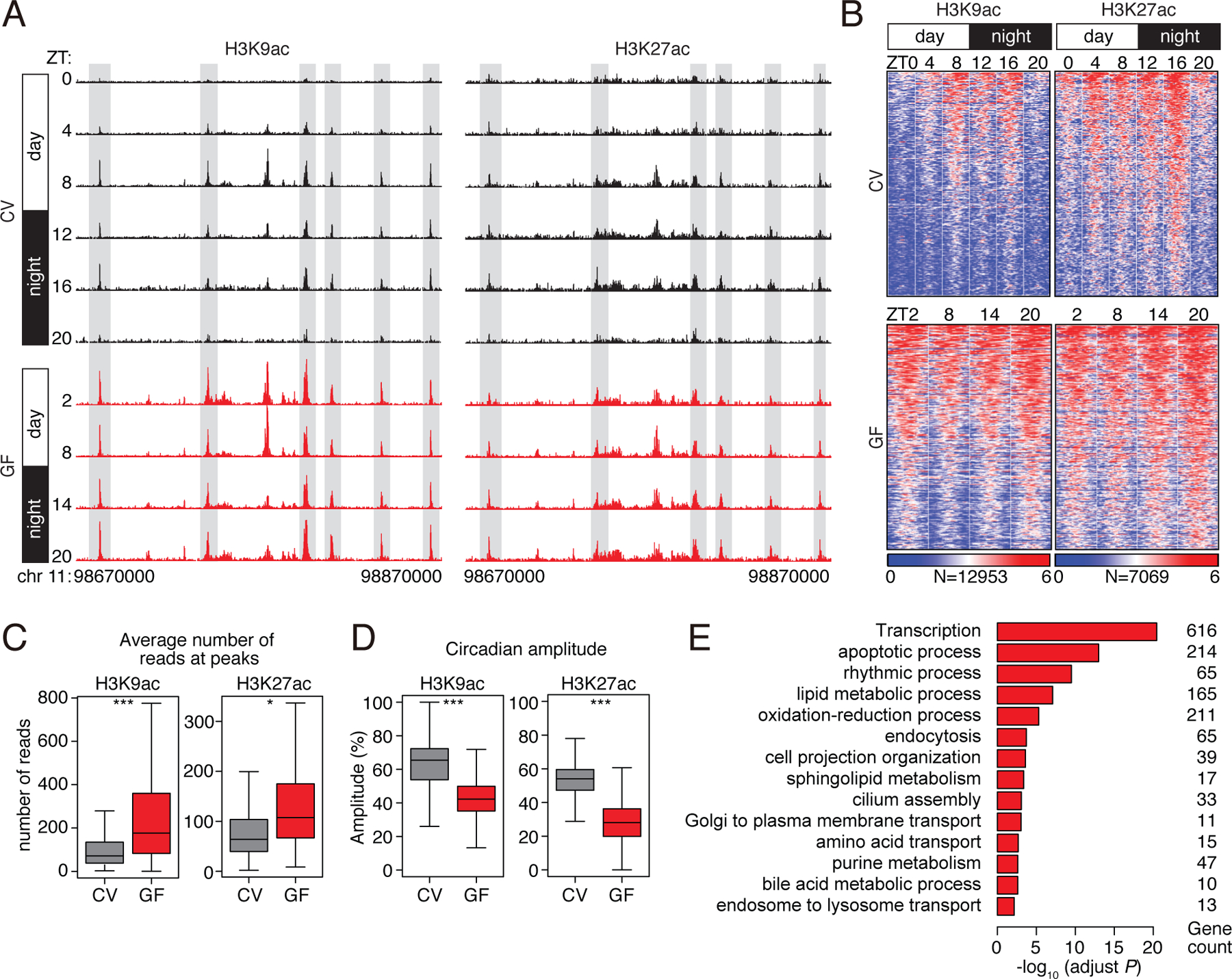
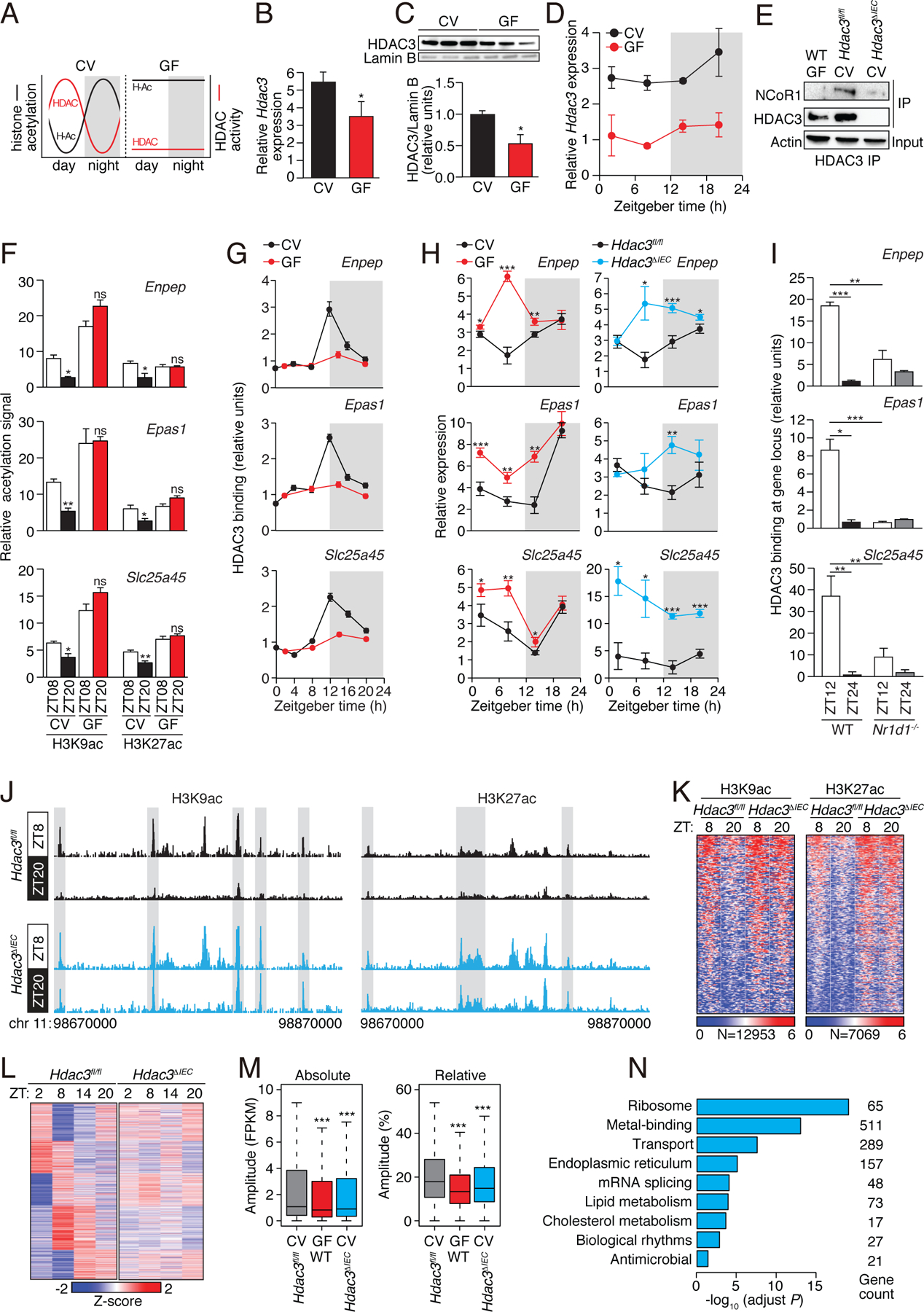
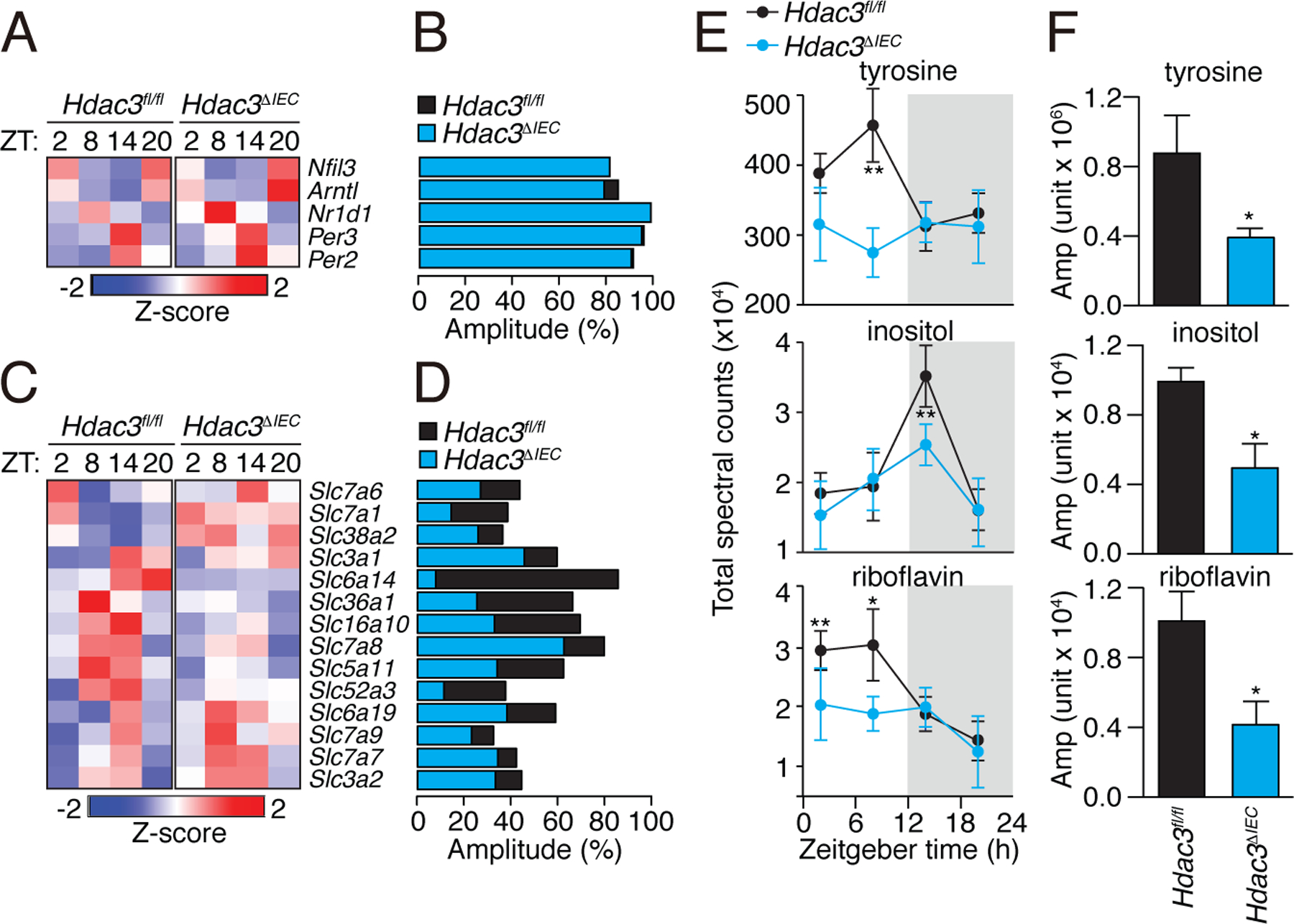
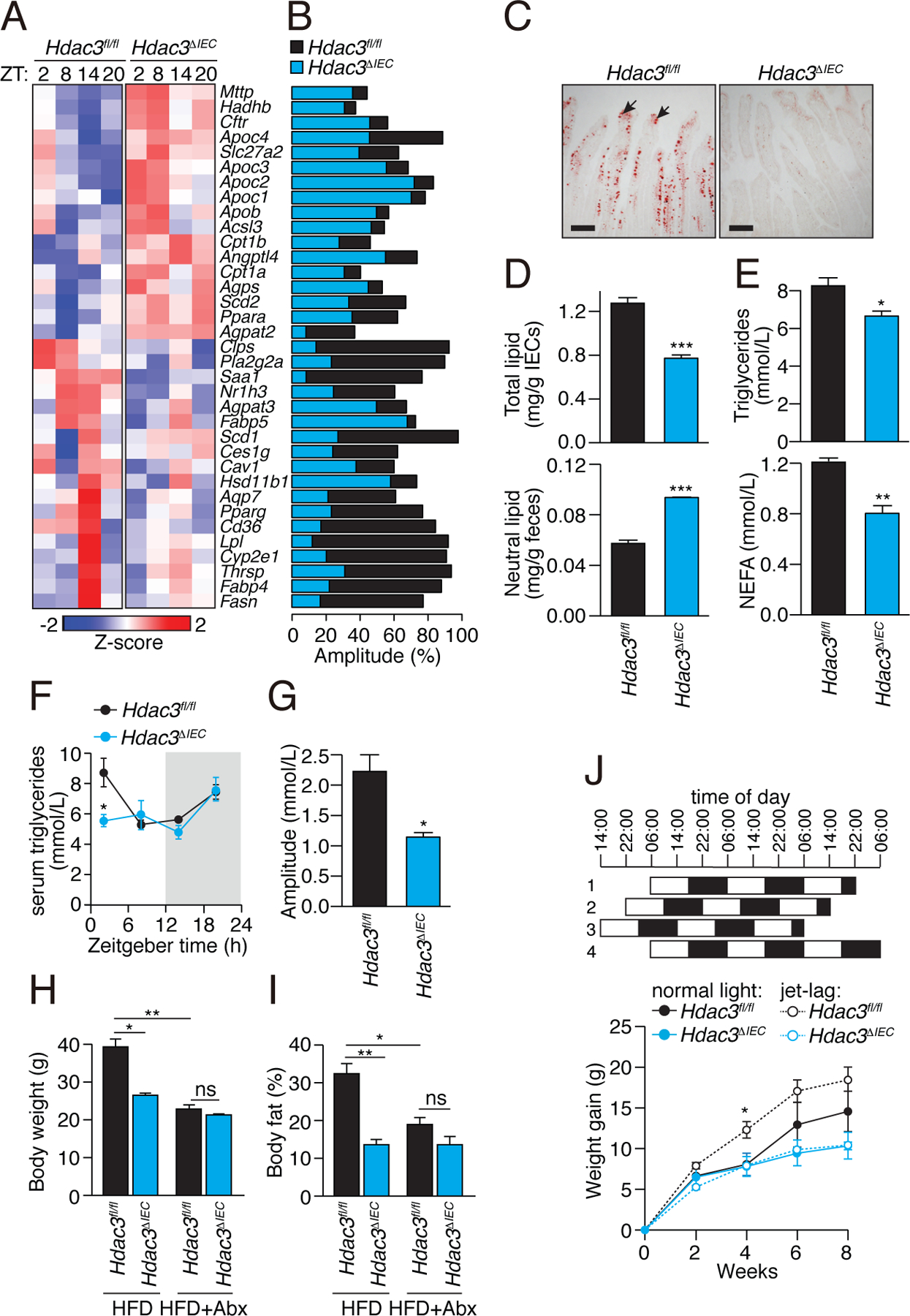
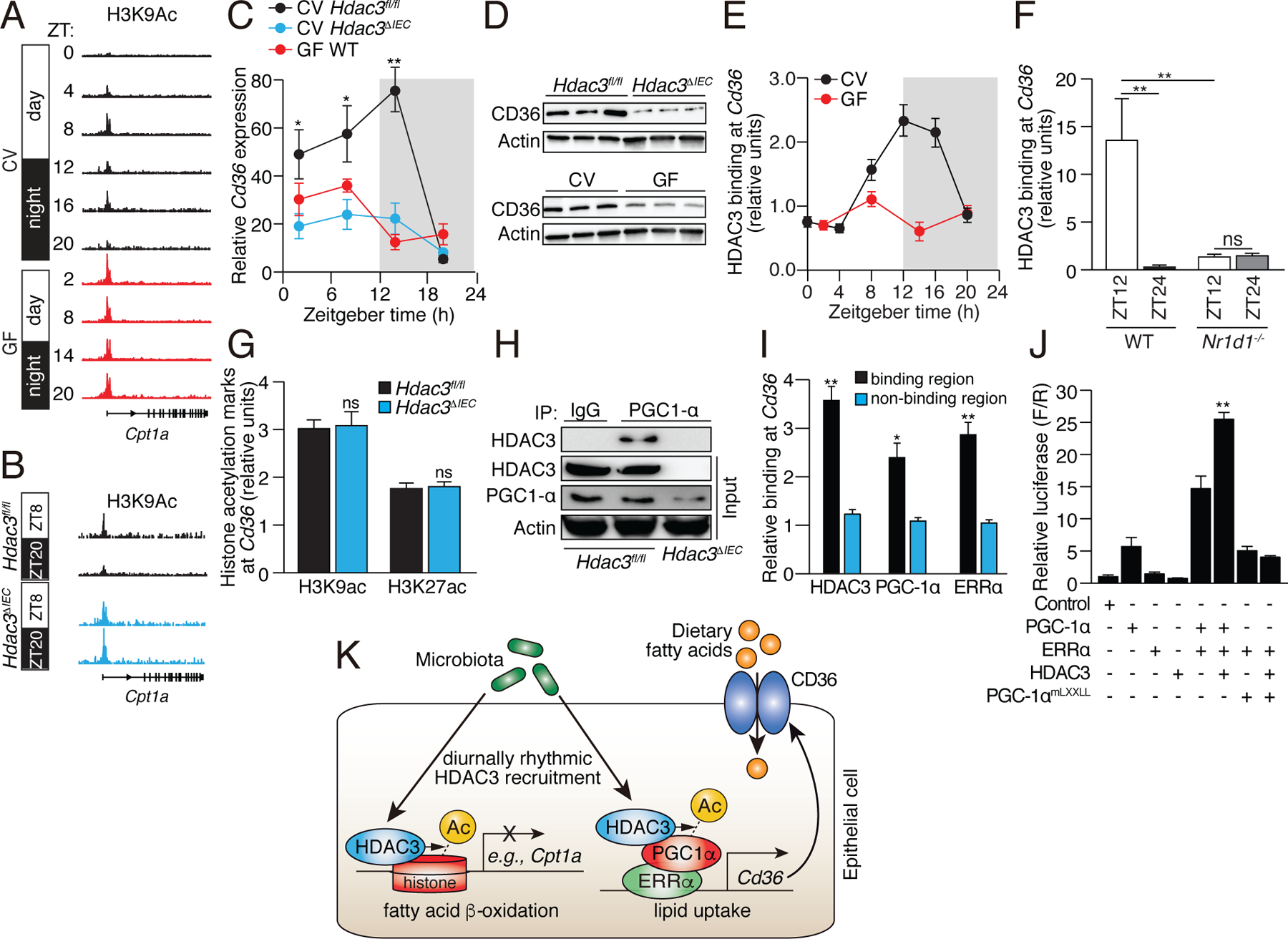
Circadian rhythmicity is a defining feature of mammalian metabolism that synchronizes metabolic processes to day-night light cycles. Here, we show that the intestinal microbiota programs diurnal metabolic rhythms in the mouse small intestine through histone deacetylase 3 (HDAC3). The microbiota induced expression of intestinal epithelial HDAC3, which was recruited rhythmically to chromatin, and produced synchronized diurnal oscillations in histone acetylation, metabolic gene expression, and nutrient uptake. HDAC3 also functioned noncanonically to coactivate estrogen-related receptor α, inducing microbiota-dependent rhythmic transcription of the lipid transporter gene Cd36 and promoting lipid absorption and diet-induced obesity. Our findings reveal that HDAC3 integrates microbial and circadian cues for regulation of diurnal metabolic rhythms and pinpoint a key mechanism by which the microbiota controls host metabolism.
Copyright © 2019 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Comment in
-
Microbial conductors.Nat Rev Microbiol. 2019 Dec;17(12):722-723. doi: 10.1038/s41579-019-0285-3. Nat Rev Microbiol. 2019. PMID: 31591501 No abstract available.
-
Microbes help to track time.Science. 2019 Sep 27;365(6460):1379-1380. doi: 10.1126/science.aaz0224. Science. 2019. PMID: 31604257 No abstract available.
References
-
- Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI, An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 444, 1027–1031 (2006). - PubMed
-
- Thaiss CA, Levy M, Korem T, Dohnalova L, Shapiro H, Jaitin DA, David E, Winter DR, Gury-BenAri M, Tatirovsky E, Tuganbaev T, Federici S, Zmora N, Zeevi D, Dori-Bachash M, Pevsner-Fischer M, Kartvelishvily E, Brandis A, Harmelin A, Shibolet O, Halpern Z, Honda K, Amit I, Segal E, Elinav E, Microbiota diurnal rhythmicity programs host transcriptome oscillations. Cell 167, 1495–1510 e1412 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
