

Calcitonin Gene-Related Peptide Negatively Regulates Alarmin-Driven Type 2 Innate Lymphoid Cell Responses
- PMID: 31604686
- PMCID: PMC7076585
- DOI: 10.1016/j.immuni.2019.09.005
Calcitonin Gene-Related Peptide Negatively Regulates Alarmin-Driven Type 2 Innate Lymphoid Cell Responses
Abstract
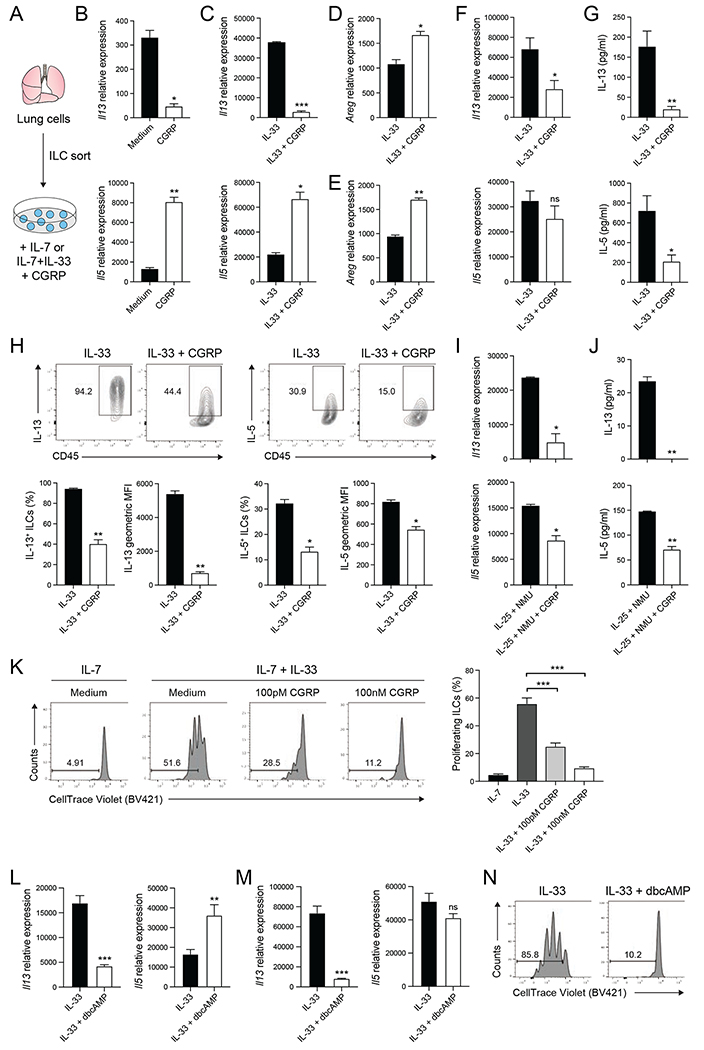
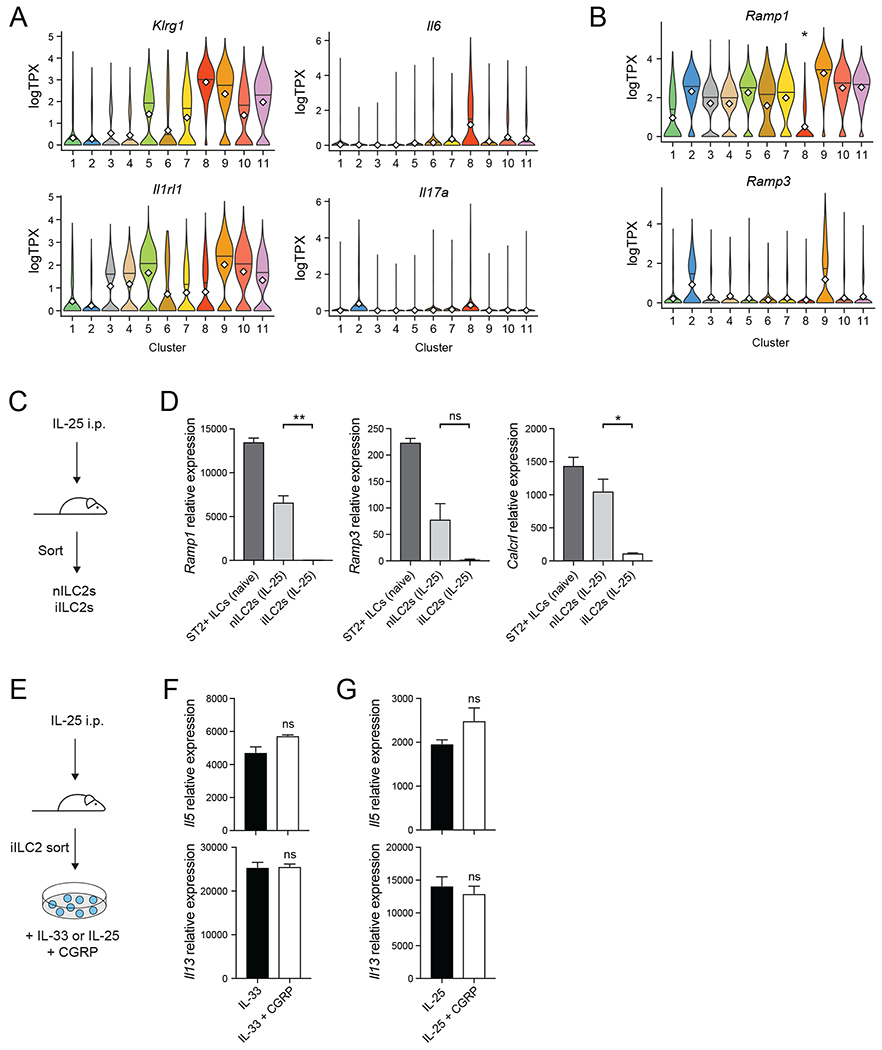
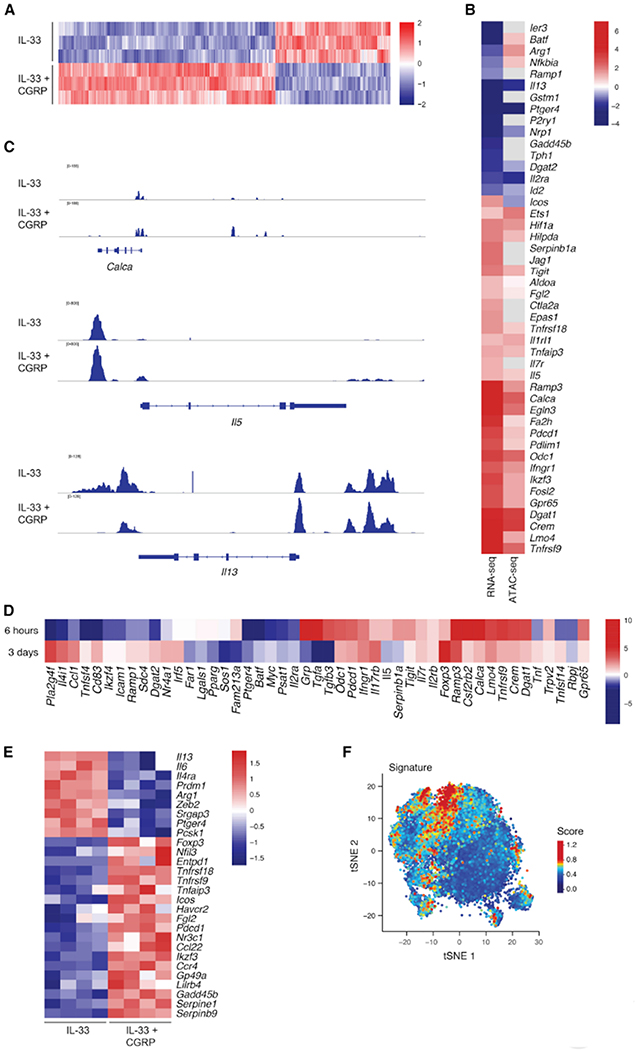
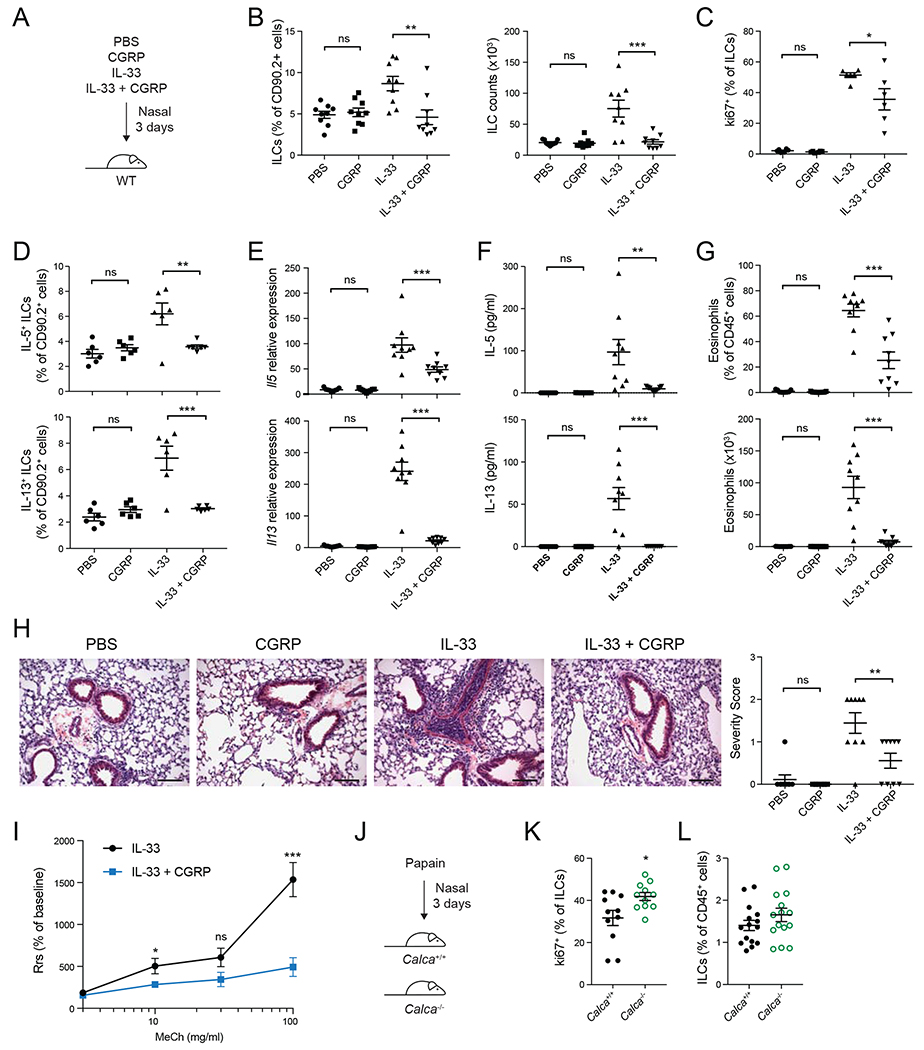
Neuroimmune interactions have emerged as critical modulators of allergic inflammation, and type 2 innate lymphoid cells (ILC2s) are an important cell type for mediating these interactions. Here, we show that ILC2s expressed both the neuropeptide calcitonin gene-related peptide (CGRP) and its receptor. CGRP potently inhibited alarmin-driven type 2 cytokine production and proliferation by lung ILC2s both in vitro and in vivo. CGRP induced marked changes in ILC2 expression programs in vivo and in vitro, attenuating alarmin-driven proliferative and effector responses. A distinct subset of ILCs scored highly for a CGRP-specific gene signature after in vivo alarmin stimulation, suggesting CGRP regulated this response. Finally, we observed increased ILC2 proliferation and type 2 cytokine production as well as exaggerated responses to alarmins in mice lacking the CGRP receptor. Together, these data indicate that endogenous CGRP is a critical negative regulator of ILC2 responses in vivo.
Keywords: CGRP; Ramp1; airway inflammation; allergic inflammation; neuro-immune interaction; neuropeptides; type 2 innate lymphoid cells.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
A.R. is an SAB member of ThermoFisher Scientific, Neogene Therapeutics, and Syros Pharmaceuticals. A.R. and V.KK. are cofounders of and equity holders in Celsius Therapeutics. A.W., P.R.B., S.J.R., A.R. and V.K.K. are co-inventors on US provisional patent application no. 62/667,381 and A.R. and R.J.X. are co-inventors on US provisional patent application no. 62/818,404, which are both filed by The Broad Institute relating to CGRP signaling modulation. All other authors have no competing financial interests.
Figures
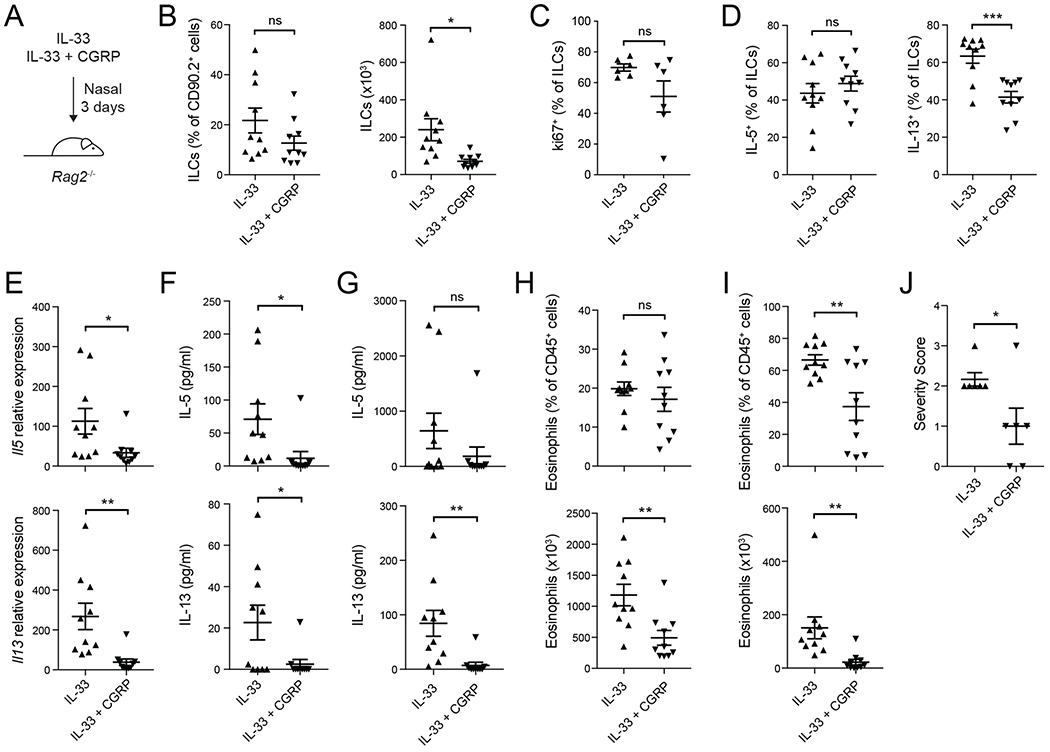
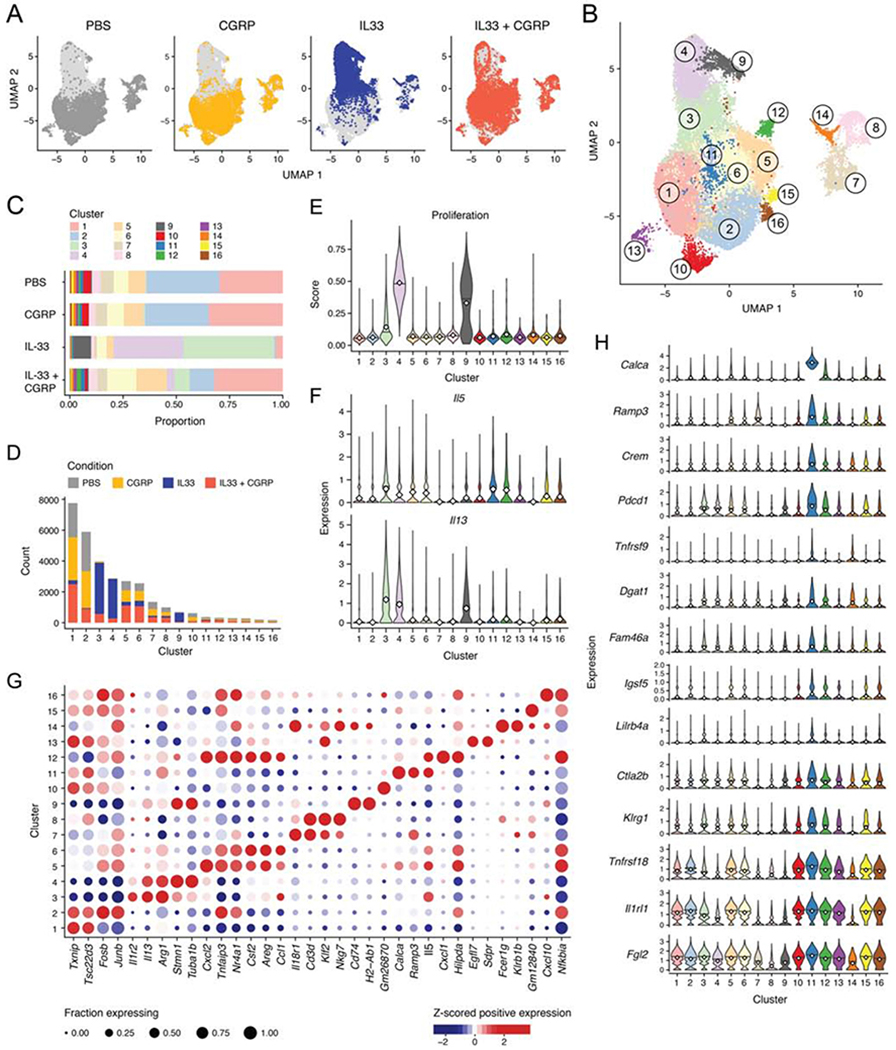
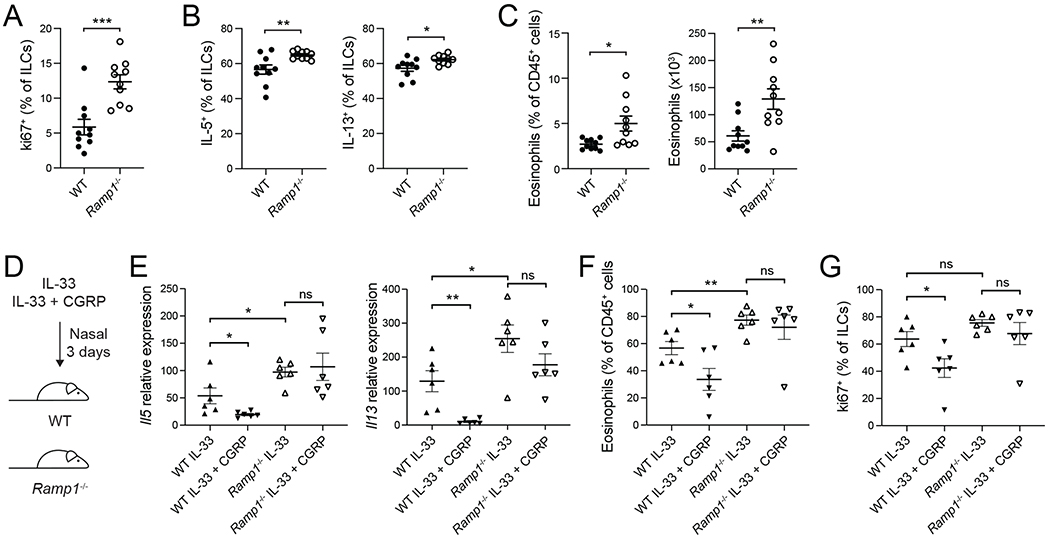
Comment in
-
The Neuropeptide CGRP Induces Bipolar Syndrome in Group 2 Innate Lymphoid Cells.Immunity. 2019 Oct 15;51(4):598-600. doi: 10.1016/j.immuni.2019.09.015. Immunity. 2019. PMID: 31618650
References
-
- Aoki-Nagase T, Nagase T, Oh-Hashi Y, Shindo T, Kurihara Y, Yamaguchi Y, Yamamoto H, Tomita T, Ohga E, Nagai R, et al. (2002). Attenuation of antigen-induced airway hyperresponsiveness in CGRP-deficient mice. Am J Physiol Lung Cell Mol Physiol 283, L963–970. - PubMed
-
- Baliu-Pique M, Jusek G, and Holzmann B (2014). Neuroimmunological communication via CGRP promotes the development of a regulatory phenotype in TLR4-stimulated macrophages. European journal of immunology 44, 3708–3716. - PubMed
-
- Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, Oukka M, Weiner HL, and Kuchroo VK (2006). Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature 441, 235–238. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
