

Pneumococci Can Become Virulent by Acquiring a New Capsule From Oral Streptococci
- PMID: 31605125
- PMCID: PMC7457184
- DOI: 10.1093/infdis/jiz456
Pneumococci Can Become Virulent by Acquiring a New Capsule From Oral Streptococci
Abstract
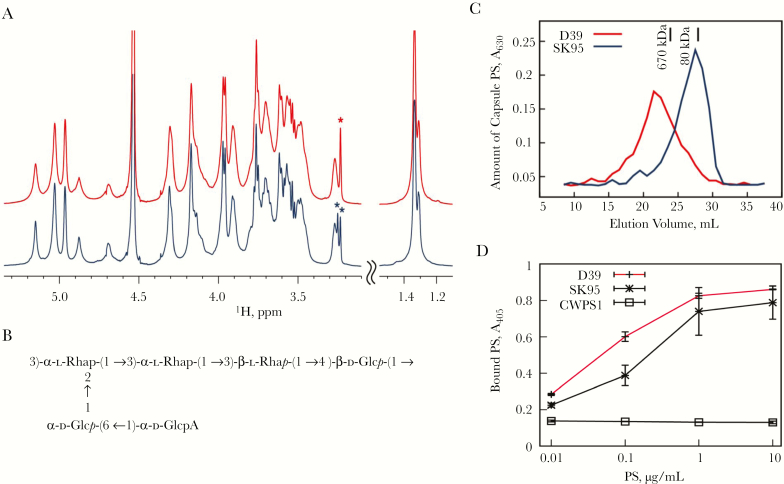
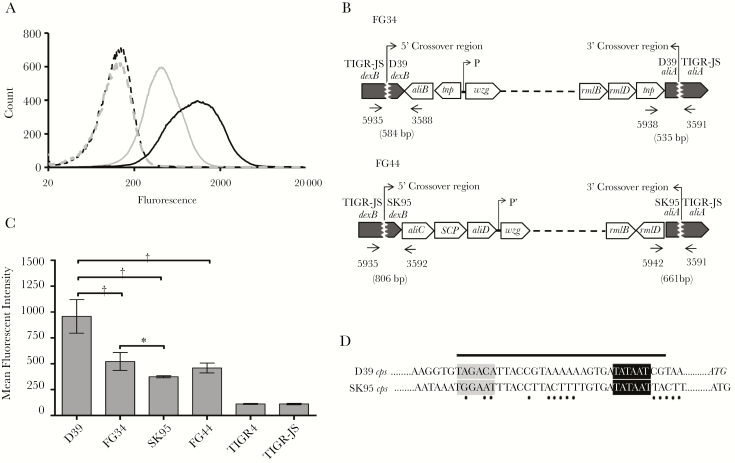
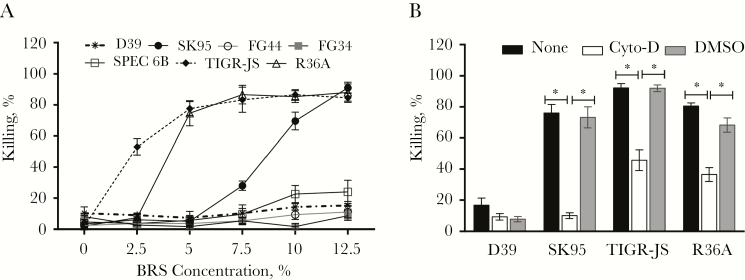
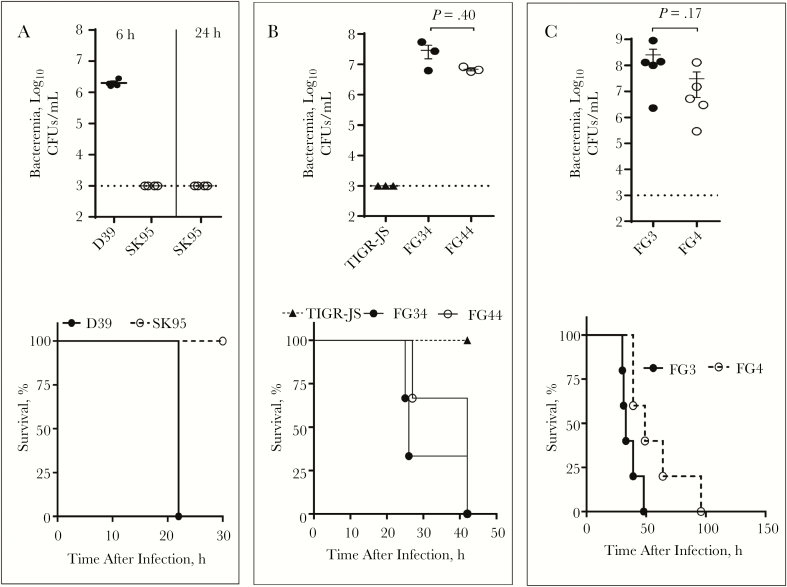
Pneumococcal conjugate vaccines have been successful, but their use has increased infections by nonvaccine serotypes. Oral streptococci often harbor capsular polysaccharide (PS) synthesis loci (cps). Although this has not been observed in nature, if pneumococcus can replace its cps with oral streptococcal cps, it may increase its serotype repertoire. In the current study, we showed that oral Streptococcus strain SK95 and pneumococcal strain D39 both produce structurally identical capsular PS, and their genetic backgrounds influence the amount of capsule production and shielding from nonspecific killing. SK95 is avirulent in a well-established in vivo mouse model. When acapsular pneumococcus was transformed with SK95 cps, the transformant became virulent and killed all mice. Thus, cps from oral Streptococcus strains can make acapsular pneumococcus virulent, and interspecies cps transfer should be considered a potential mechanism of serotype replacement. Our findings, along with publications from the US Centers for Disease Control and Prevention, highlight potential limitations of the 2013 World Health Organization criterion for studying pneumococcal serotypes carried without isolating bacteria. We show that an oral streptococcal strain, SK95, and a pneumococcal strain, D39, both produce chemically identical capsular PS. We also show that transferring SK95 cps into noncapsulated, avirulent pneumococcus gave it the capacity for virulence in a mouse model.
Keywords: pneumococcus; serotype; vaccine.
© The Author(s) 2019. Published by Oxford University Press for the Infectious Diseases Society of America. All rights reserved. For permissions, e-mail: journals.permissions@oup.com.
Figures
Comment in
-
Potential Epidemiologic and Historical Implications of Capsular Serotypes Shared by Pneumococci and Their Nonpneumococcal Relatives.J Infect Dis. 2020 Jul 6;222(3):343-346. doi: 10.1093/infdis/jiz457. J Infect Dis. 2020. PMID: 31605123 No abstract available.
References
-
- Drijkoningen JJC, Rohde GGU. Pneumococcal infection in adults: burden of disease. Clin Microbiol Infect 2014; 20:45–51. - PubMed
-
- World Health Organization. Pneumococcal conjugate vaccine for childhood immunization—WHO position paper. Wkly Epidemiol Rec 2007; 82:93–104. - PubMed
-
- Diawara I, Zerouali K, Katfy K, et al. . Invasive pneumococcal disease among children younger than 5 years of age before and after introduction of pneumococcal conjugate vaccine in Casablanca, Morocco. Int J Infect Dis 2015; 40:95–101. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
