

TCF-1-Centered Transcriptional Network Drives an Effector versus Exhausted CD8 T Cell-Fate Decision
- PMID: 31606264
- PMCID: PMC6943829
- DOI: 10.1016/j.immuni.2019.09.013
TCF-1-Centered Transcriptional Network Drives an Effector versus Exhausted CD8 T Cell-Fate Decision
Abstract
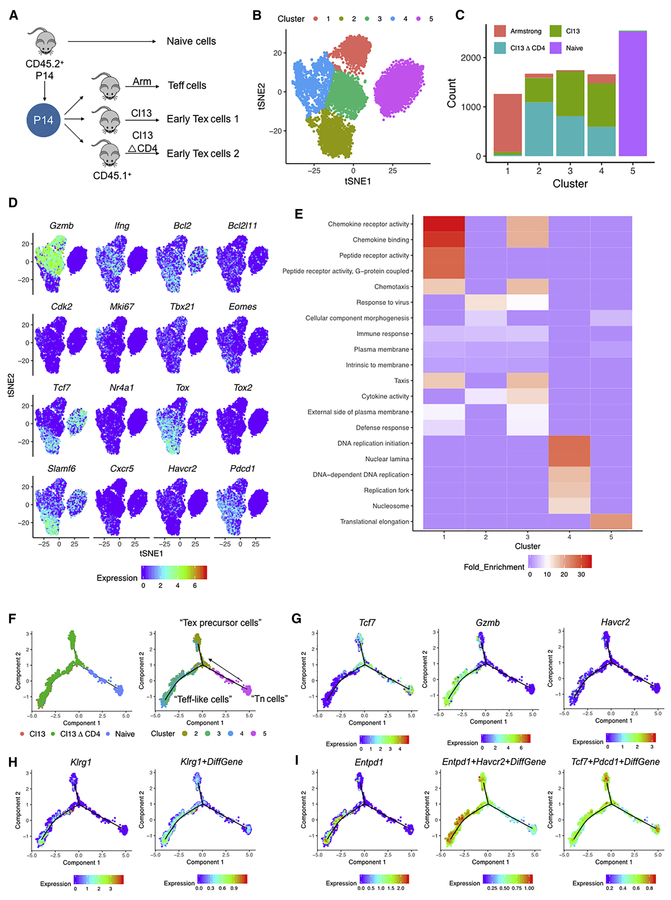
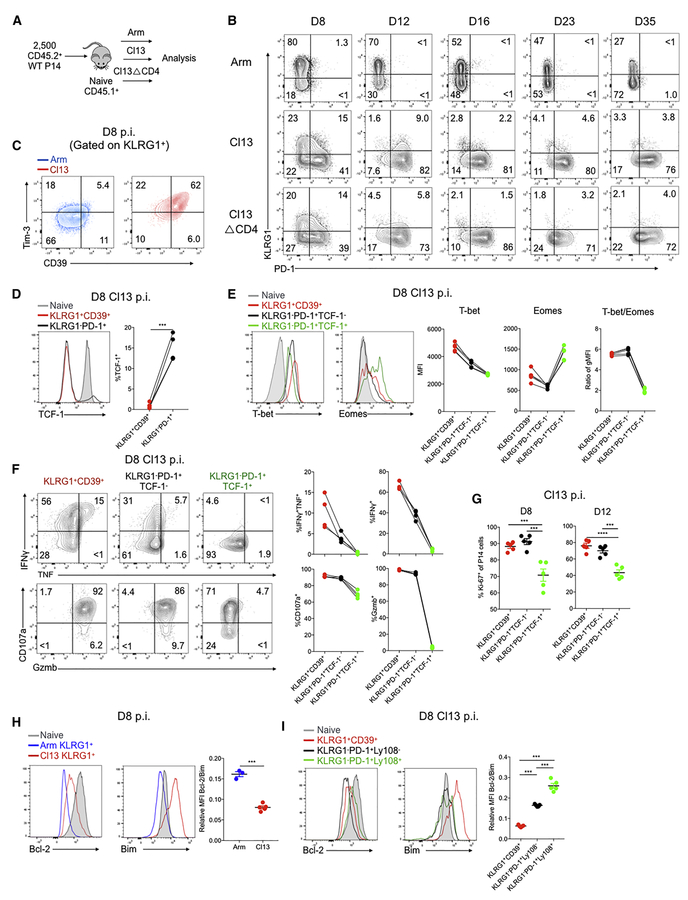
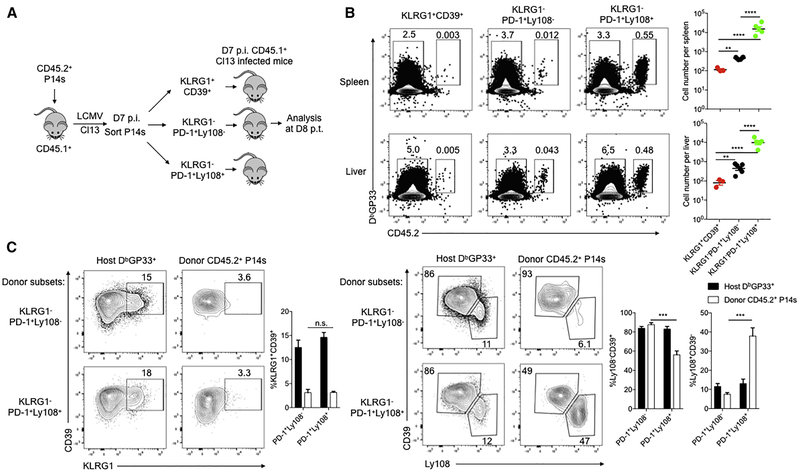
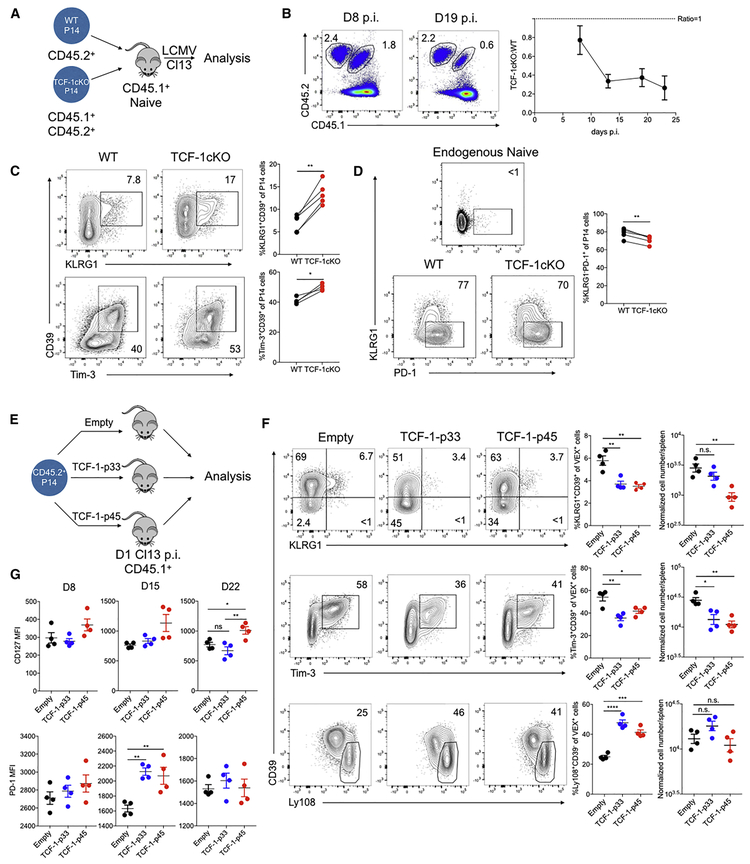
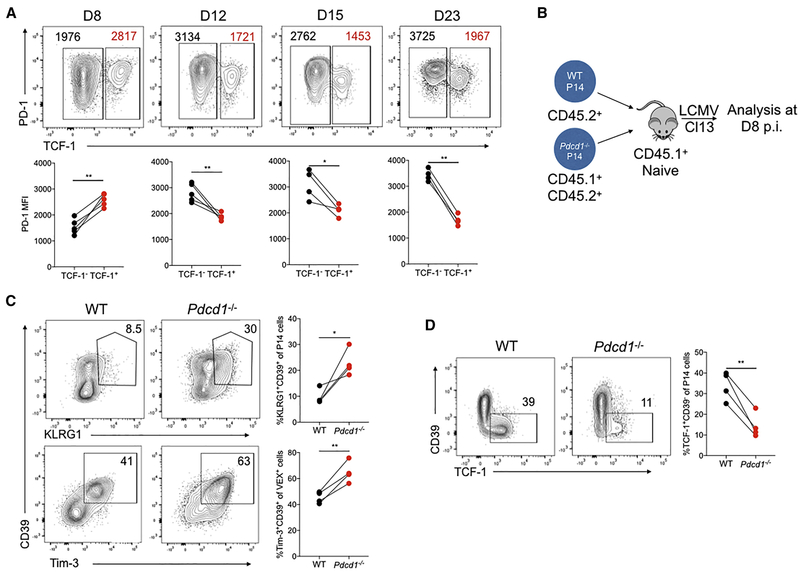
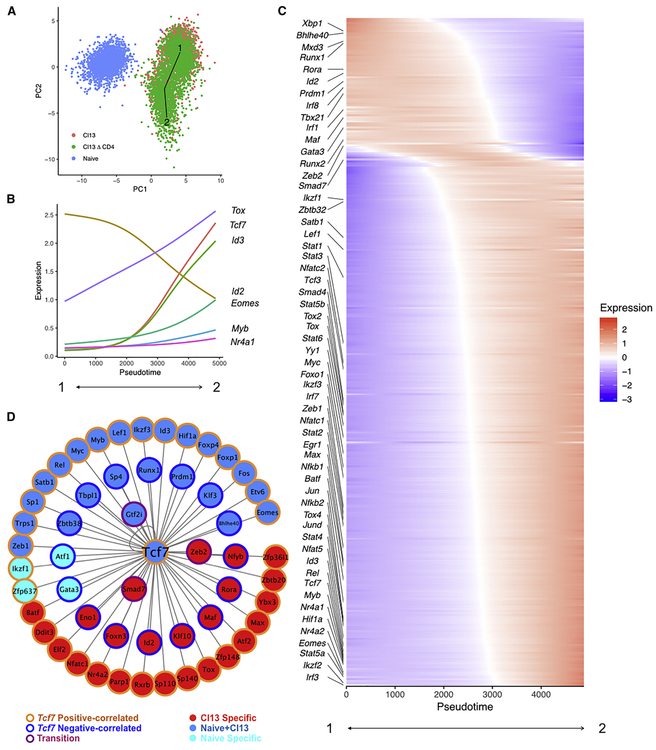
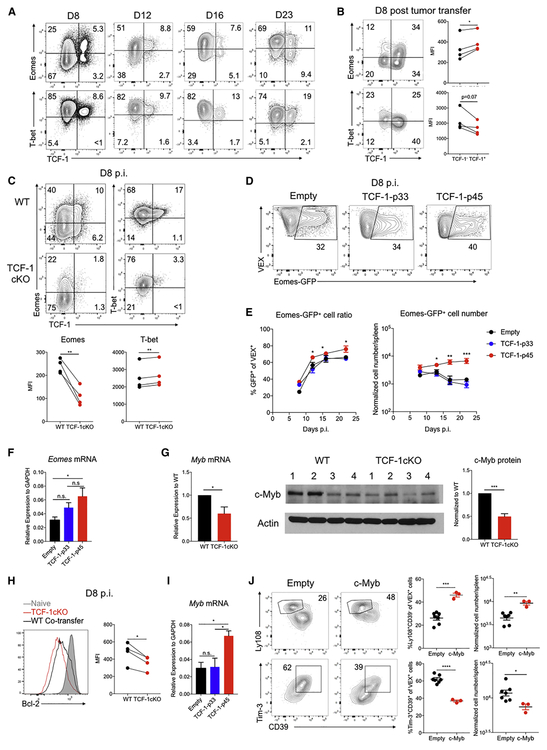
TCF-1 is a key transcription factor in progenitor exhausted CD8 T cells (Tex). Moreover, this Tex cell subset mediates responses to PD-1 checkpoint pathway blockade. However, the role of the transcription factor TCF-1 in early fate decisions and initial generation of Tex cells is unclear. Single-cell RNA sequencing (scRNA-seq) and lineage tracing identified a TCF-1+Ly108+PD-1+ CD8 T cell population that seeds development of mature Tex cells early during chronic infection. TCF-1 mediated the bifurcation between divergent fates, repressing development of terminal KLRG1Hi effectors while fostering KLRG1Lo Tex precursor cells, and PD-1 stabilized this TCF-1+ Tex precursor cell pool. TCF-1 mediated a T-bet-to-Eomes transcription factor transition in Tex precursors by promoting Eomes expression and drove c-Myb expression that controlled Bcl-2 and survival. These data define a role for TCF-1 in early-fate-bifurcation-driving Tex precursor cells and also identify PD-1 as a protector of this early TCF-1 subset.
Keywords: CD8 T cell exhaustion; PD-1; cancer; chronic infection; exhaustion; immunotherapy; transcriptional circuit.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
E.J.W. has consulting agreements with and/or is on the scientific advisory board for Merck, Roche, Pieris, Elstar, and Surface Oncology. E.J.W. is a founder of Surface Oncology and Arsenal Biosciences. E.J.W. has a patent licensing agreement on the PD-1 pathway with Roche/Genentech.
Figures
Comment in
-
Back to the Future: Effector Fate during T Cell Exhaustion.Immunity. 2019 Dec 17;51(6):970-972. doi: 10.1016/j.immuni.2019.11.007. Immunity. 2019. PMID: 31851902
References
-
- Adams WC, Chen Y-H, Kratchmarov R, Yen B, Nish SA, Lin W-HW, Rothman NJ, Luchsinger LL, Klein U, Busslinger M, Rathmell JC, Snoeck H-W, Reiner SL, 2016. Anabolism-Associated Mitochondrial Stasis Driving Lymphocyte Differentiation over Self-Renewal. Cell Reports 17, 3142–3152. doi: 10.1016/j.celrep.2016.11.065 - DOI - PMC - PubMed
-
- Alfei F, Kanev K, Hofmann M, Wu M, Ghoneim HE, Roelli P, Utzschneider DT, Hösslin, von M, Cullen JG, Fan Y, Eisenberg V, Wohlleber D, Steiger K, Merkler D, Delorenzi M, Knolle PA, Cohen CJ, Thimme R, Youngblood B, Zehn D, 2019. TOX reinforces the phenotype and longevity of exhausted T cells in chronic viral infection. Nature 2017 545:7652 1. doi: 10.1038/s41586-019-1326-9 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 HG009518/HG/NHGRI NIH HHS/United States
- U19 AI117950/AI/NIAID NIH HHS/United States
- T32 CA009140/CA/NCI NIH HHS/United States
- F99 CA234842/CA/NCI NIH HHS/United States
- K08 AI114852/AI/NIAID NIH HHS/United States
- U19 AI082630/AI/NIAID NIH HHS/United States
- P01 AI112521/AI/NIAID NIH HHS/United States
- R01 AI115712/AI/NIAID NIH HHS/United States
- K00 CA234842/CA/NCI NIH HHS/United States
- P01 AI108545/AI/NIAID NIH HHS/United States
- R01 AI105343/AI/NIAID NIH HHS/United States
- P01 CA210944/CA/NCI NIH HHS/United States
- K08 AI136660/AI/NIAID NIH HHS/United States
- R01 CA078831/CA/NCI NIH HHS/United States
- R01 HL145754/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
