

Altered RNA Processing in Cancer Pathogenesis and Therapy
- PMID: 31611195
- PMCID: PMC6825565
- DOI: 10.1158/2159-8290.CD-19-0399
Altered RNA Processing in Cancer Pathogenesis and Therapy
Abstract
Major advances in our understanding of cancer pathogenesis and therapy have come from efforts to catalog genomic alterations in cancer. A growing number of large-scale genomic studies have uncovered mutations that drive cancer by perturbing cotranscriptional and post-transcriptional regulation of gene expression. These include alterations that affect each phase of RNA processing, including splicing, transport, editing, and decay of messenger RNA. The discovery of these events illuminates a number of novel therapeutic vulnerabilities generated by aberrant RNA processing in cancer, several of which have progressed to clinical development. SIGNIFICANCE: There is increased recognition that genetic alterations affecting RNA splicing and polyadenylation are common in cancer and may generate novel therapeutic opportunities. Such mutations may occur within an individual gene or in RNA processing factors themselves, thereby influencing splicing of many downstream target genes. This review discusses the biological impact of these mutations on tumorigenesis and the therapeutic approaches targeting cells bearing these mutations.
©2019 American Association for Cancer Research.
Figures
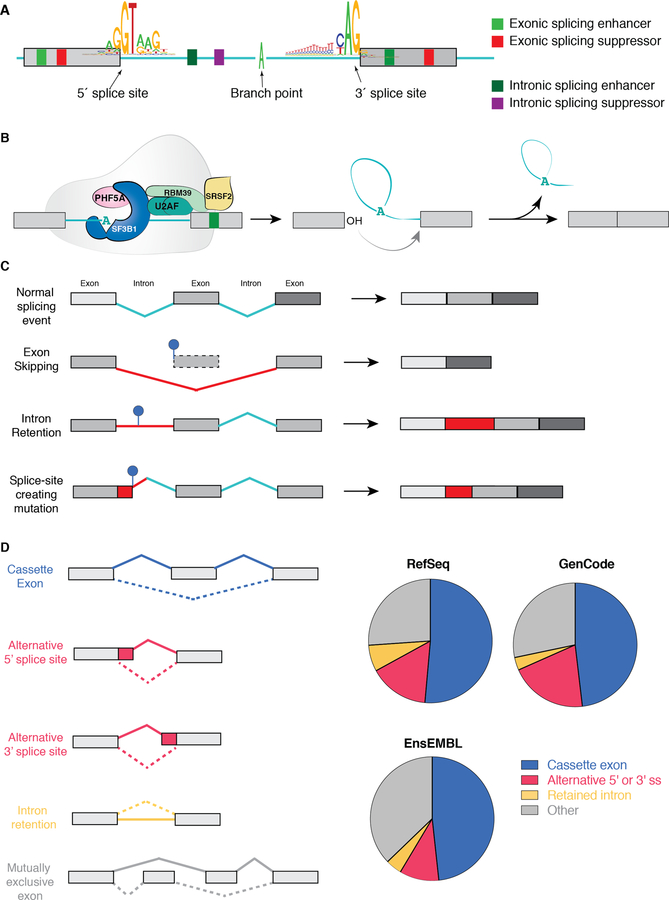
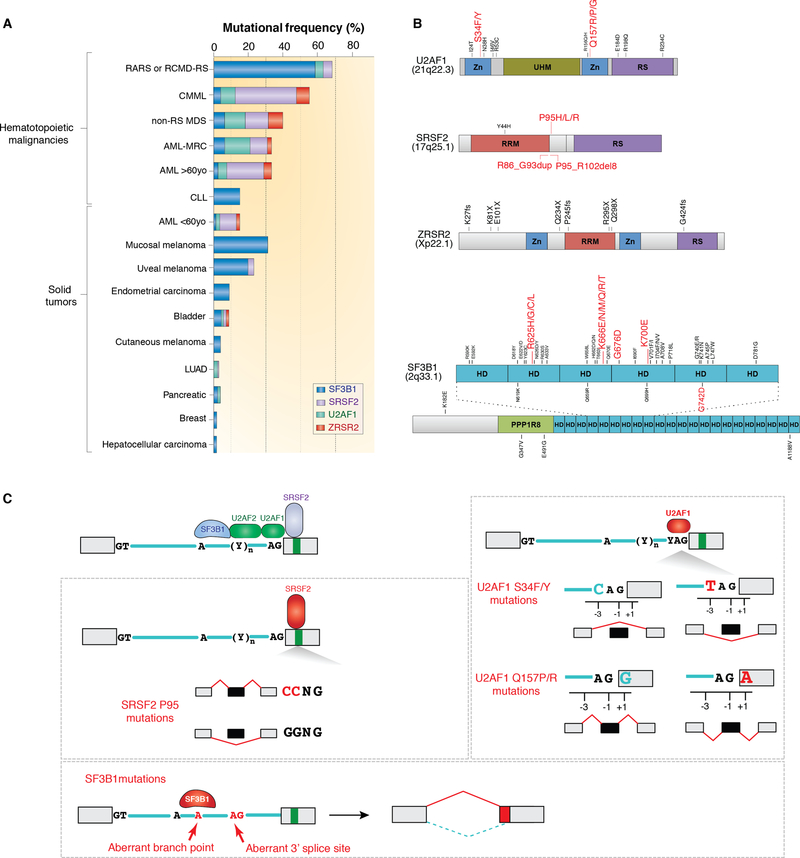
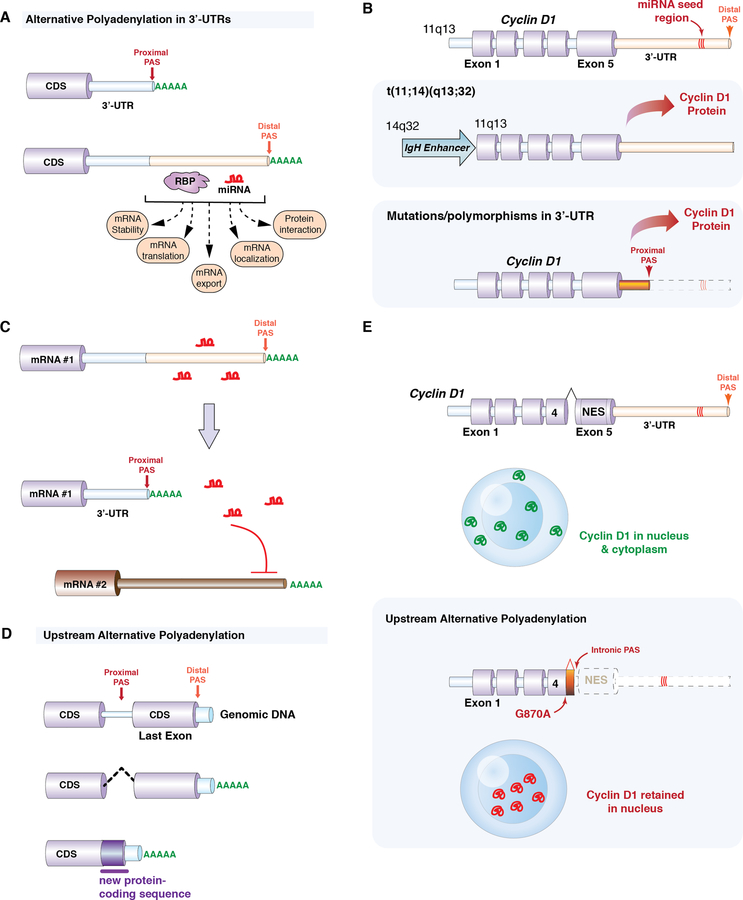
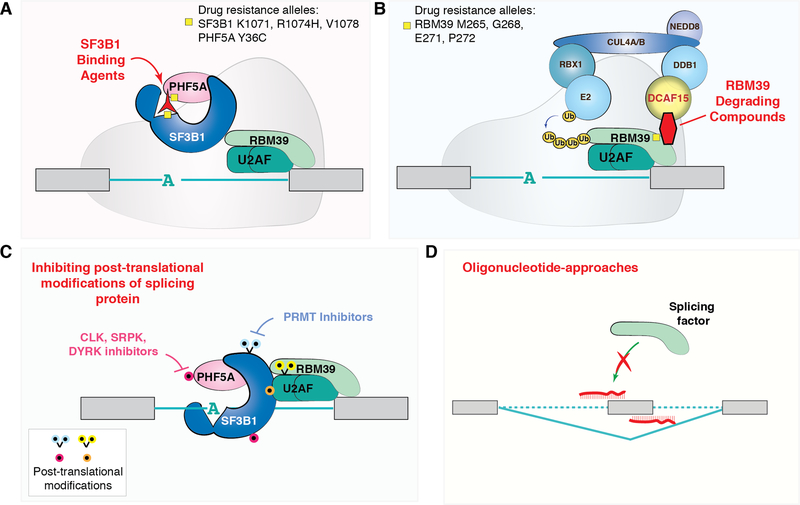
References
-
- Vu LP, Cheng Y, Kharas MG. The Biology of m(6)A RNA Methylation in Normal and Malignant Hematopoiesis. Cancer discovery 2019;9:25–33. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
