

Distinct molecular etiologies of male and female hepatocellular carcinoma
- PMID: 31615477
- PMCID: PMC6794913
- DOI: 10.1186/s12885-019-6167-2
Distinct molecular etiologies of male and female hepatocellular carcinoma
Abstract
Background: Sex-differences in cancer occurrence and mortality are evident across tumor types; men exhibit higher rates of incidence and often poorer responses to treatment. Targeted approaches to the treatment of tumors that account for these sex-differences require the characterization and understanding of the fundamental biological mechanisms that differentiate them. Hepatocellular Carcinoma (HCC) is the second leading cause of cancer death worldwide, with the incidence rapidly rising. HCC exhibits a male-bias in occurrence and mortality, but previous studies have failed to explore the sex-specific dysregulation of gene expression in HCC.
Methods: Here, we characterize the sex-shared and sex-specific regulatory changes in HCC tumors in the TCGA LIHC cohort using combined and sex-stratified differential expression and eQTL analyses.
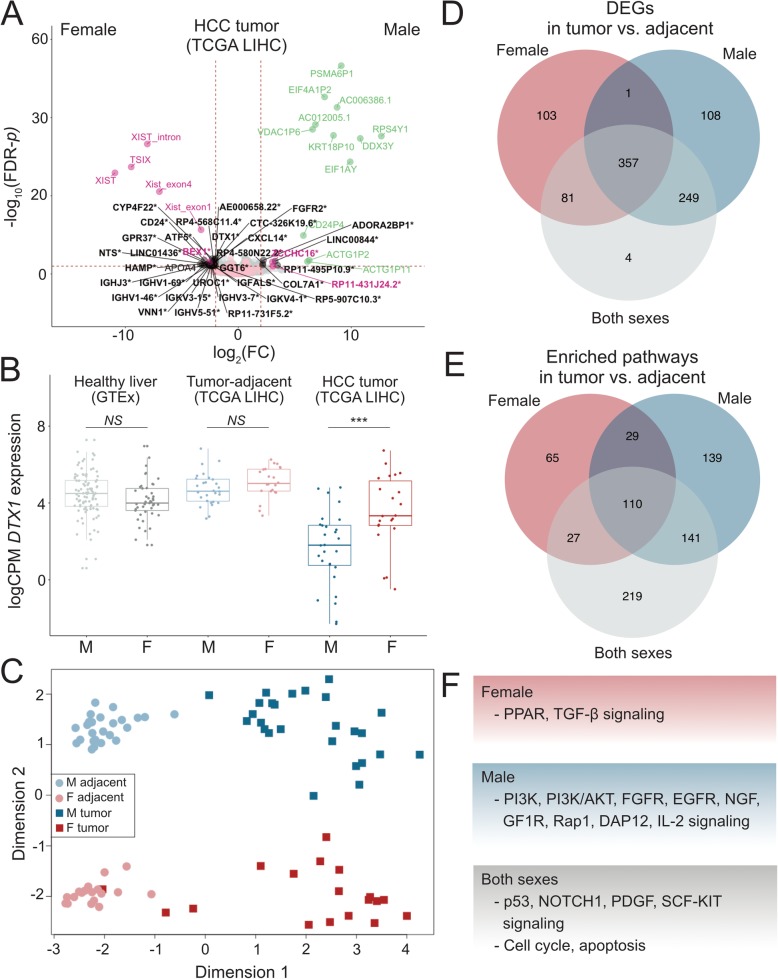
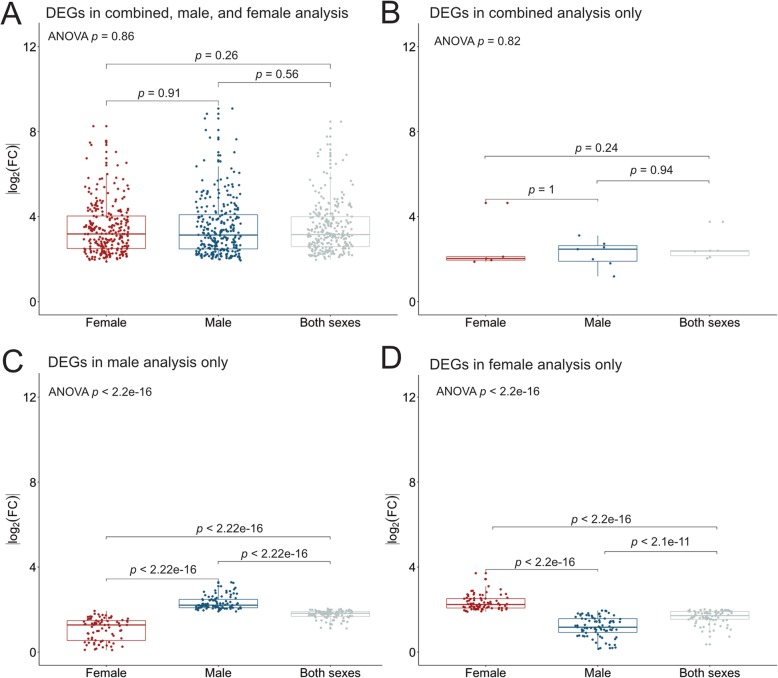
Results: By using a sex-specific differential expression analysis of tumor and tumor-adjacent samples, we uncovered etiologically relevant genes and pathways differentiating male and female HCC. While both sexes exhibited activation of pathways related to apoptosis and cell cycle, males and females differed in the activation of several signaling pathways, with females showing PPAR pathway enrichment while males showed PI3K, PI3K/AKT, FGFR, EGFR, NGF, GF1R, Rap1, DAP12, and IL-2 signaling pathway enrichment. Using eQTL analyses, we discovered germline variants with differential effects on tumor gene expression between the sexes. 24.3% of the discovered eQTLs exhibit differential effects between the sexes, illustrating the substantial role of sex in modifying the effects of eQTLs in HCC. The genes that showed sex-specific dysregulation in tumors and those that harbored a sex-specific eQTL converge in clinically relevant pathways, suggesting that the molecular etiologies of male and female HCC are partially driven by differential genetic effects on gene expression.
Conclusions: Sex-stratified analyses detect sex-specific molecular etiologies of HCC. Overall, our results provide new insight into the role of inherited genetic regulation of transcription in modulating sex-differences in HCC etiology and provide a framework for future studies on sex-biased cancers.
Keywords: Gene expression; HCC; Hepatocellular carcinoma; Sex; Sex as a biological variable; eQTL.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
-
- Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018. 10.3322/caac.21492. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
