

Developmental changes in plasticity, synaptic, glia, and connectivity protein levels in rat basolateral amygdala
- PMID: 31615855
- PMCID: PMC6796789
- DOI: 10.1101/lm.049866.119
Developmental changes in plasticity, synaptic, glia, and connectivity protein levels in rat basolateral amygdala
Abstract
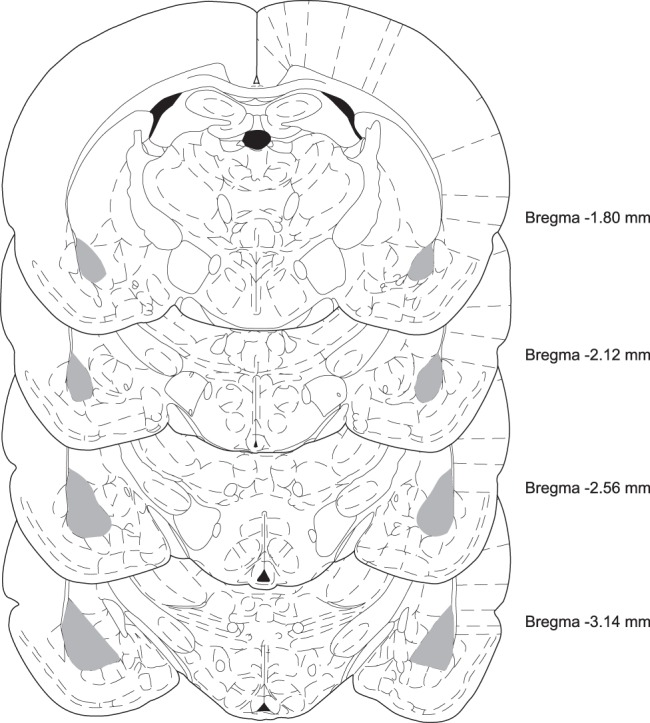
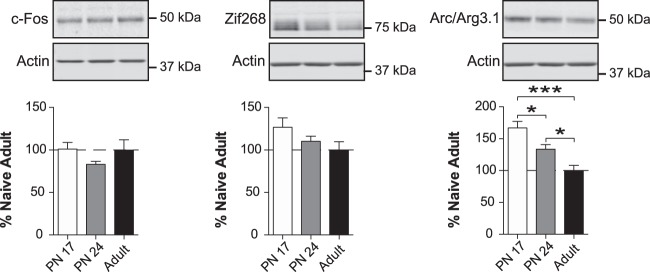
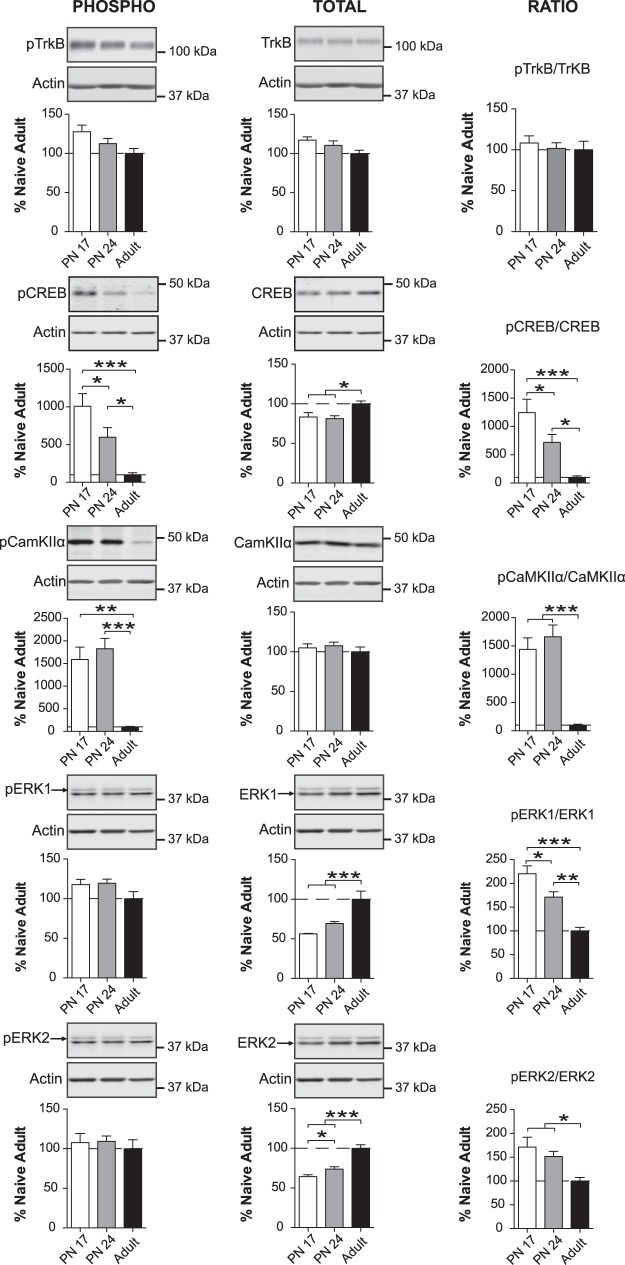
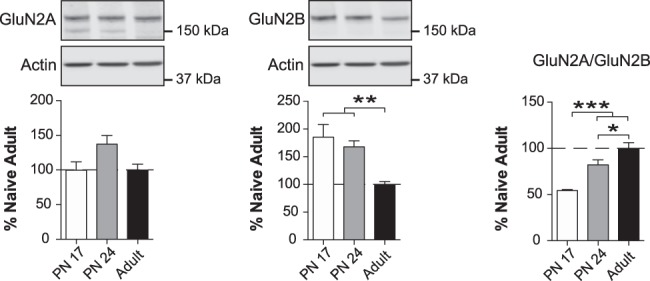
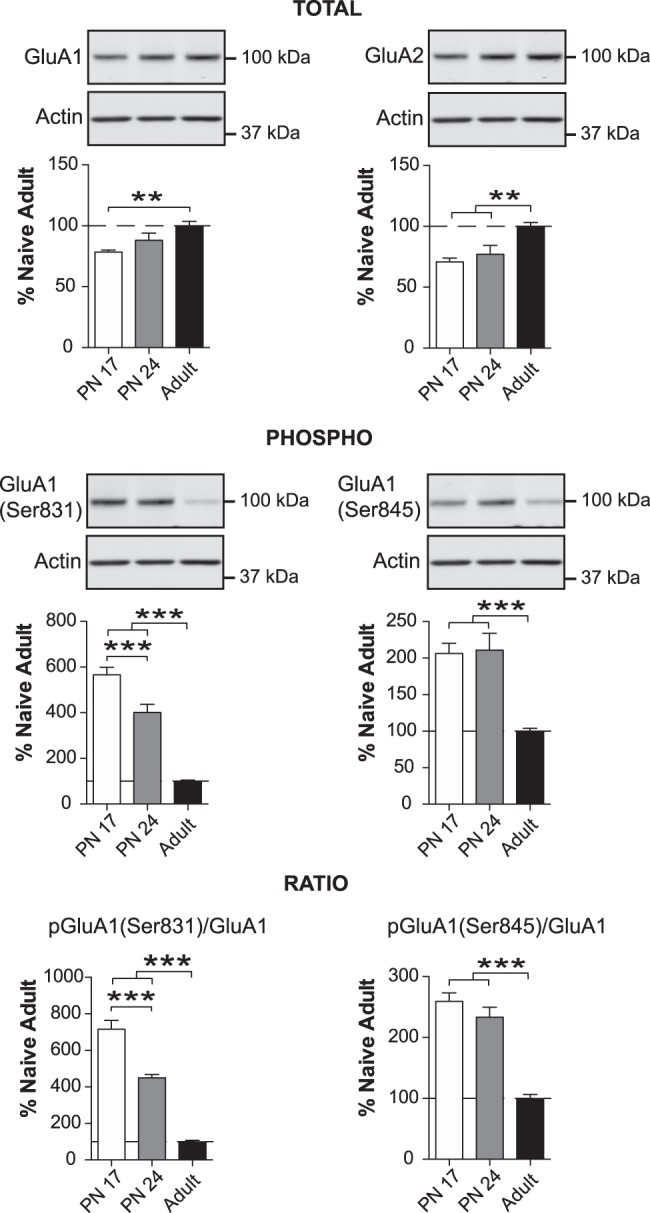
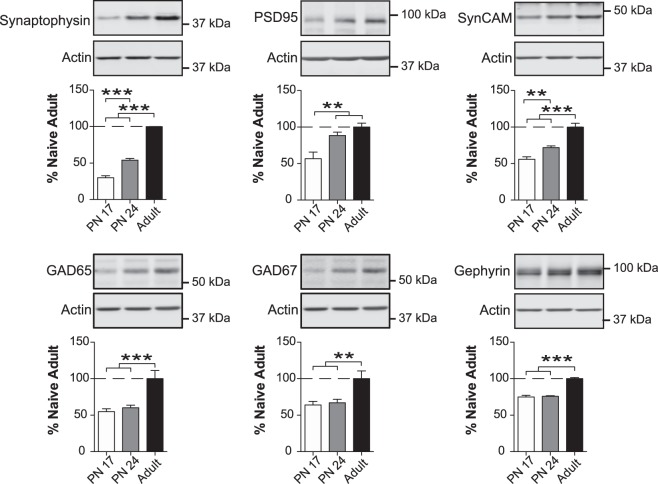
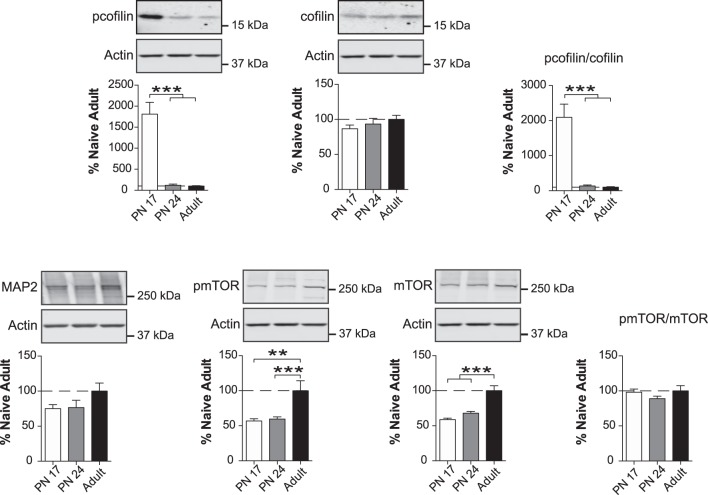
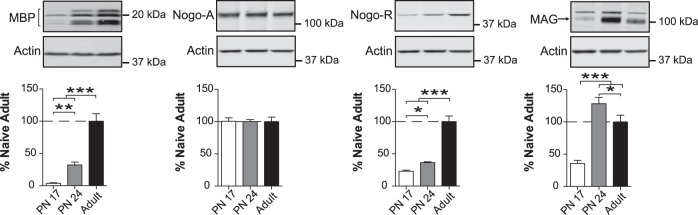
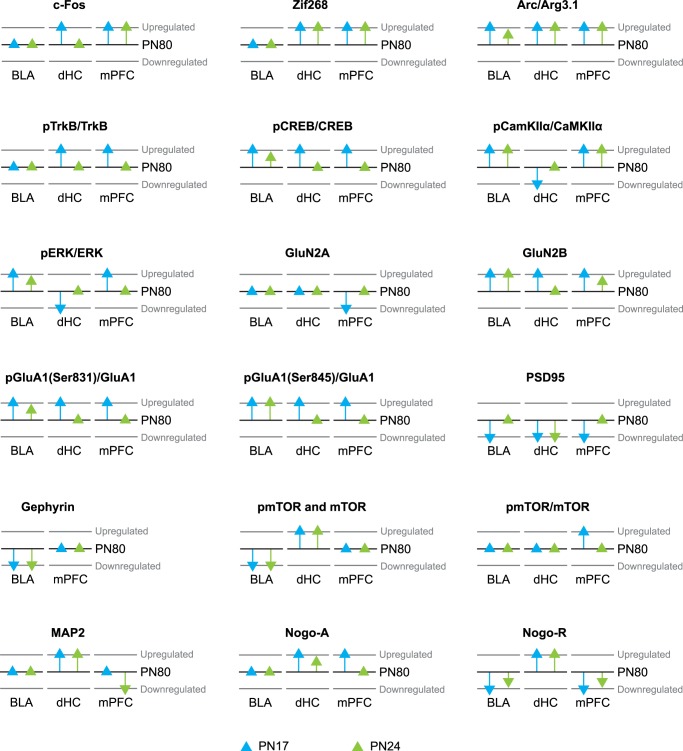
The basolateral complex of amygdala (BLA) processes emotionally arousing aversive and rewarding experiences. The BLA is critical for acquisition and storage of threat-based memories and the modulation of the consolidation of arousing explicit memories, that is, the memories that are encoded and stored by the medial temporal lobe. In addition, in conjunction with the medial prefrontal cortex (mPFC), the BLA plays an important role in fear memory extinction. The BLA develops relatively early in life, but little is known about the molecular changes that accompany its development. Here, we quantified relative basal expression levels of sets of plasticity, synaptic, glia, and connectivity proteins in the rat BLA at various developmental ages: postnatal day 17 (PN17, infants), PN24 (juveniles), and PN80 (young adults). We found that the levels of activation markers of brain plasticity, including phosphorylation of CREB at Ser133, CamKIIα at Thr286, pERK1/pERK2 at Thr202/Tyr204, and GluA1 at Ser831 and Ser845, were significantly higher in infant and juvenile compared with adult brain. In contrast, age increase was accompanied by a significant augmentation in the levels of proteins that mark synaptogenesis and synapse maturation, such as synaptophysin, PSD95, SynCAM, GAD65, GAD67, and GluN2A/GluN2B ratio. Finally, we observed significant age-associated changes in structural markers, including MAP2, MBP, and MAG, suggesting that the structural connectivity of the BLA increases over time. The biological differences in the BLA between developmental ages compared with adulthood suggest the need for caution in extrapolating conclusions based on BLA-related brain plasticity and behavioral studies conducted at different developmental stages.
© 2019 Bessières et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous