

Kinetic and structural evidence that Asp-678 plays multiple roles in catalysis by the quinoprotein glycine oxidase
- PMID: 31615898
- PMCID: PMC6873188
- DOI: 10.1074/jbc.RA119.011255
Kinetic and structural evidence that Asp-678 plays multiple roles in catalysis by the quinoprotein glycine oxidase
Abstract
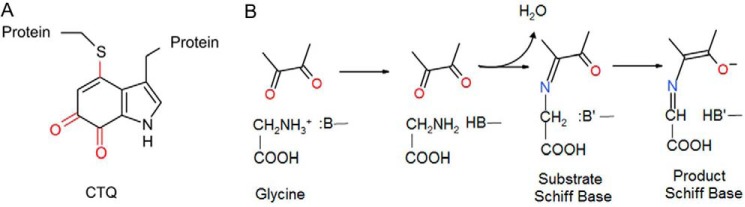
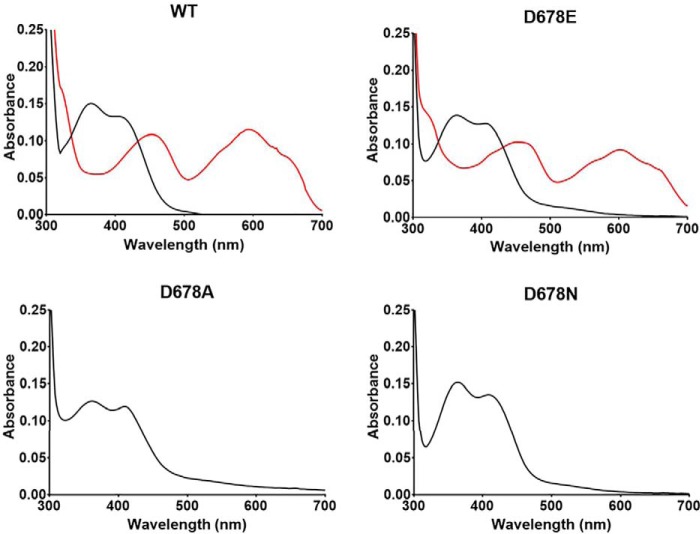
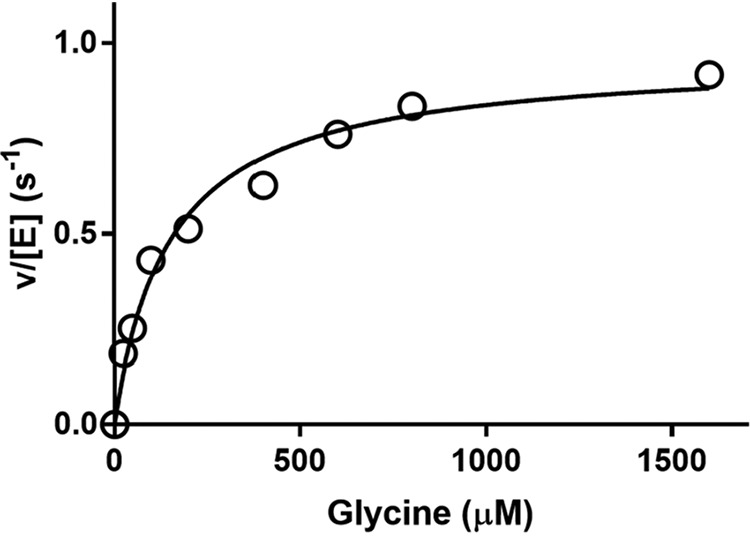
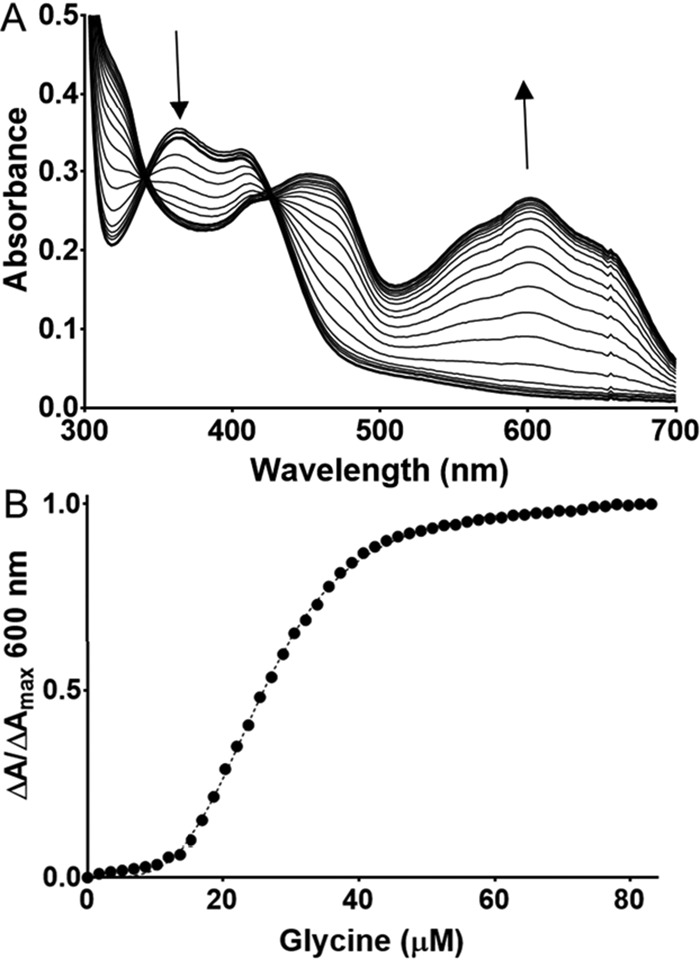
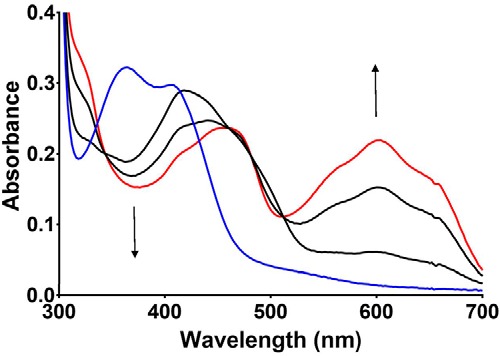
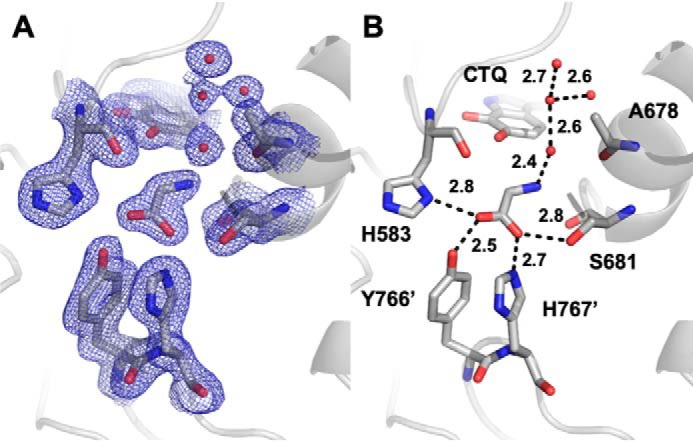
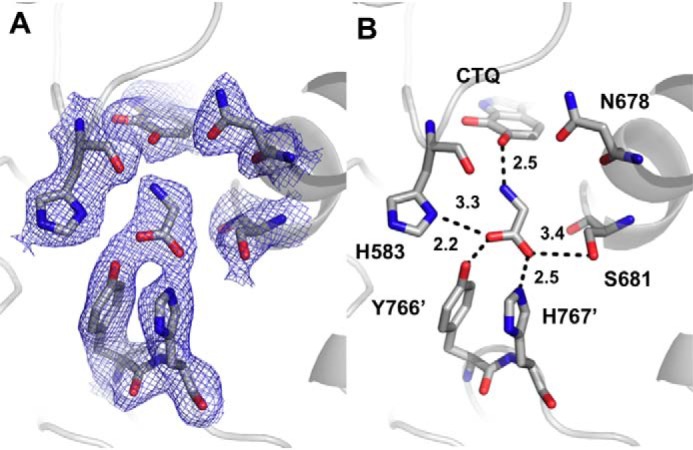
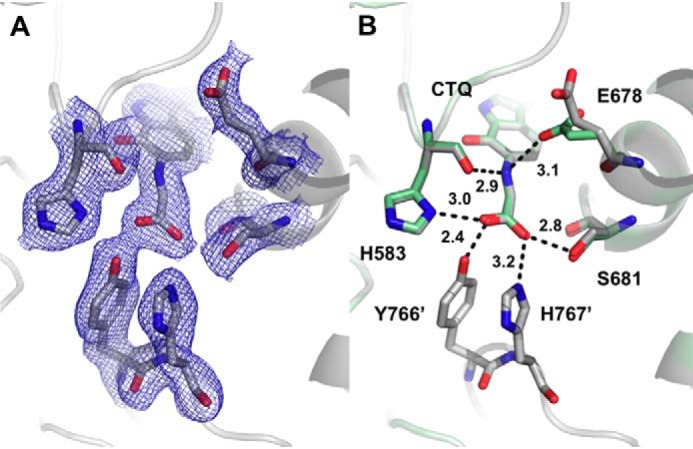
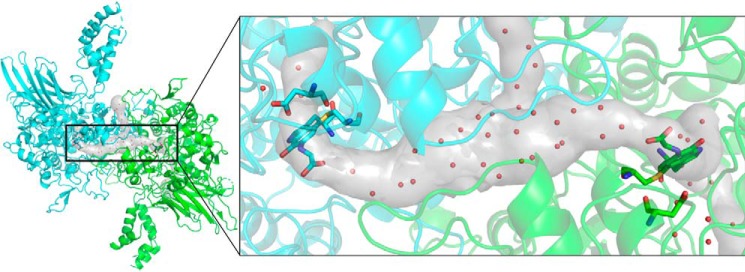
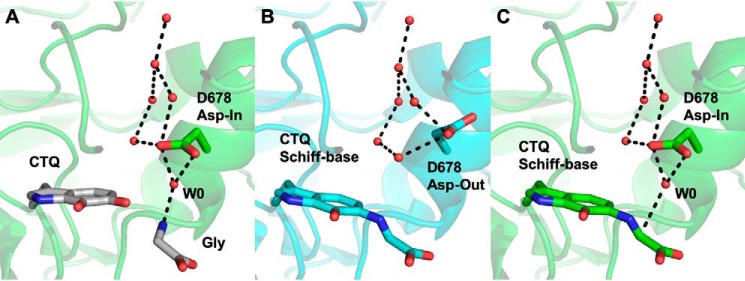
PlGoxA from Pseudoalteromonas luteoviolacea is a glycine oxidase that utilizes a protein-derived cysteine tryptophylquinone (CTQ) cofactor. A notable feature of its catalytic mechanism is that it forms a stable product-reduced CTQ adduct that is not hydrolyzed in the absence of O2 Asp-678 resides near the quinone moiety of PlGoxA, and an Asp is structurally conserved in this position in all tryptophylquinone enzymes. In those other enzymes, mutation of that Asp results in no or negligible CTQ formation. In this study, mutation of Asp-678 in PlGoxA did not abolish CTQ formation. This allowed, for the first time, studying the role of this residue in catalysis. D678A and D678N substitutions yielded enzyme variants with CTQ, which did not react with glycine, although glycine was present in the crystal structures in the active site. D678E PlGoxA was active but exhibited a much slower kcat This mutation altered the kinetic mechanism of the reductive half-reaction such that one could observe a previously undetected reactive intermediate, an initial substrate-oxidized CTQ adduct, which converted to the product-reduced CTQ adduct. These results indicate that Asp-678 is involved in the initial deprotonation of the amino group of glycine, enabling nucleophilic attack of CTQ, as well as the deprotonation of the substrate-oxidized CTQ adduct, which is coupled to CTQ reduction. The structures also suggest that Asp-678 is acting as a proton relay that directs these protons to a water channel that connects the active sites on the subunits of this homotetrameric enzyme.
Keywords: GoxA; LodA-like protein; acid-base catalysis; allostery; cysteine tryptophylquinone (CTQ); enzyme kinetics; enzyme mechanism; glycine oxidase; oxidase; protein structure; proton transfer; proton transport; quinone; quinoprotein; water channel.
© 2019 Mamounis et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Andreo-Vidal A., Mamounis K. J., Sehanobish E., Avalos D., Campillo-Brocal J. C., Sanchez-Amat A., Yukl E. T., and Davidson V. L. (2018) Structure and enzymatic properties of an unusual cysteine tryptophylquinone-dependent glycine oxidase from Pseudoalteromonas luteoviolacea. Biochemistry 57, 1155–1165 10.1021/acs.biochem.8b00009 - DOI - PMC - PubMed
-
- Chacón-Verdú M. D., Campillo-Brocal J. C., Lucas-Elío P., Davidson V. L., and Sánchez-Amat A. (2015) Characterization of recombinant biosynthetic precursors of the cysteine tryptophylquinone cofactors of l-lysine-ϵ-oxidase and glycine oxidase from Marinomonas mediterranea. Biochim. Biophys. Acta 1854, 1123–1131 10.1016/j.bbapap.2014.12.018 - DOI - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
