

Oxidative stress-mediated TXNIP loss causes RPE dysfunction
- PMID: 31615975
- PMCID: PMC6802648
- DOI: 10.1038/s12276-019-0327-y
Oxidative stress-mediated TXNIP loss causes RPE dysfunction
Abstract
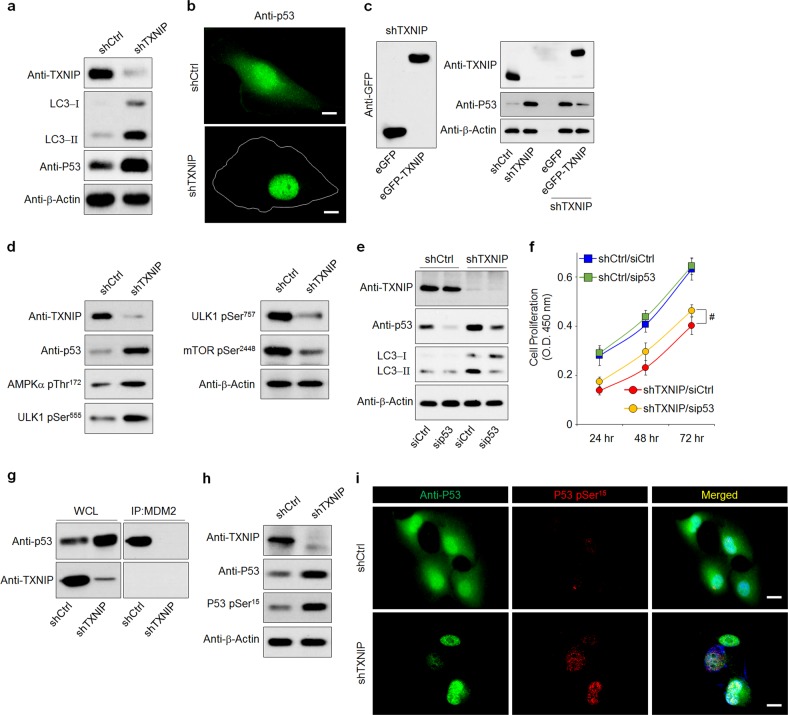
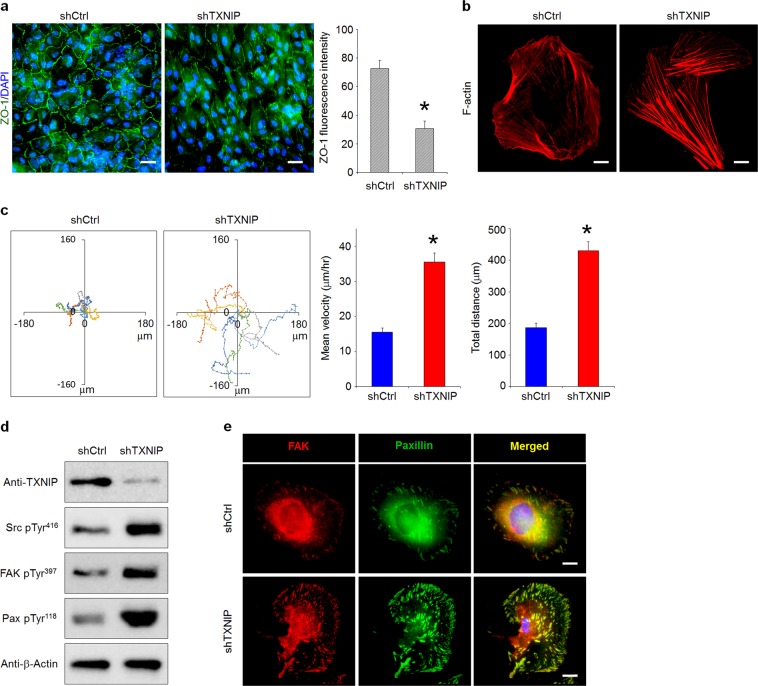
The disruption of the retinal pigment epithelium (RPE), for example, through oxidative damage, is a common factor underlying age-related macular degeneration (AMD). Aberrant autophagy also contributes to AMD pathology, as autophagy maintains RPE homeostasis to ensure blood-retinal barrier (BRB) integrity and protect photoreceptors. Thioredoxin-interacting protein (TXNIP) promotes cellular oxidative stress by inhibiting thioredoxin reducing capacity and is in turn inversely regulated by reactive oxygen species levels; however, its role in oxidative stress-induced RPE cell dysfunction and the mechanistic link between TXNIP and autophagy are largely unknown. Here, we observed that TXNIP expression was rapidly downregulated in RPE cells under oxidative stress and that RPE cell proliferation was decreased. TXNIP knockdown demonstrated that the suppression of proliferation resulted from TXNIP depletion-induced autophagic flux, causing increased p53 activation via nuclear localization, which in turn enhanced AMPK phosphorylation and activation. Moreover, TXNIP downregulation further negatively impacted BRB integrity by disrupting RPE cell tight junctions and enhancing cell motility by phosphorylating, and thereby activating, Src kinase. Finally, we also revealed that TXNIP knockdown upregulated HIF-1α, leading to the enhanced secretion of VEGF from RPE cells and the stimulation of angiogenesis in cocultured human retinal microvascular endothelial cells. This suggests that the exposure of RPE cells to sustained oxidative stress may promote choroidal neovascularization, another AMD pathology. Together, these findings reveal three distinct mechanisms by which TXNIP downregulation disrupts RPE cell function and thereby exacerbates AMD pathogenesis. Accordingly, reinforcing or restoring BRB integrity by targeting TXNIP may serve as an effective therapeutic strategy for preventing or attenuating photoreceptor damage in AMD.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures



References
-
- Nowak JZ. Age-related macular degeneration (AMD): pathogenesis and therapy. Pharmacol. Rep. 2006;58:353–363. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
