

Pathogenesis of hypertension in a mouse model for human CLCN2 related hyperaldosteronism
- PMID: 31615979
- PMCID: PMC6794291
- DOI: 10.1038/s41467-019-12113-9
Pathogenesis of hypertension in a mouse model for human CLCN2 related hyperaldosteronism
Abstract
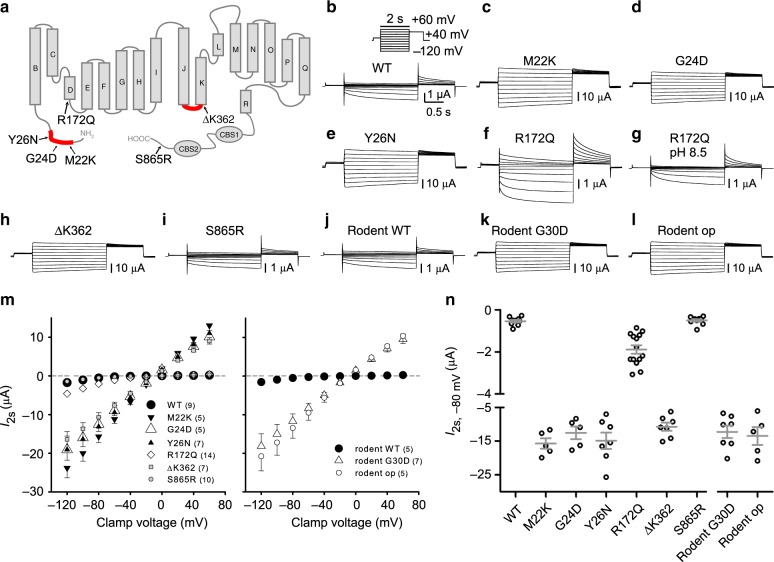
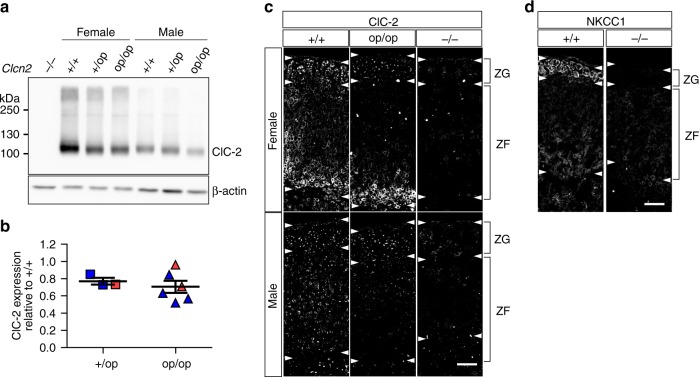
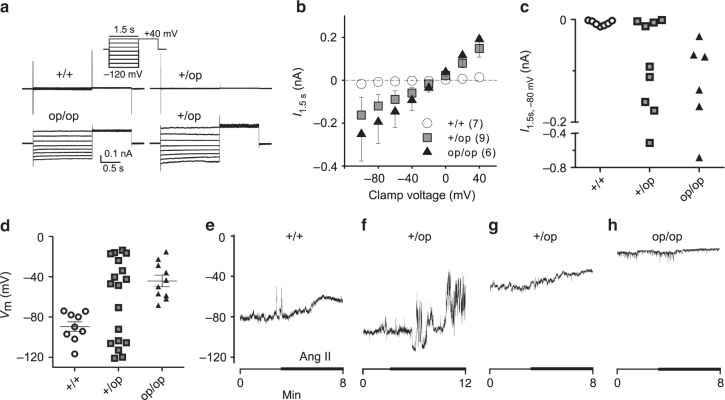
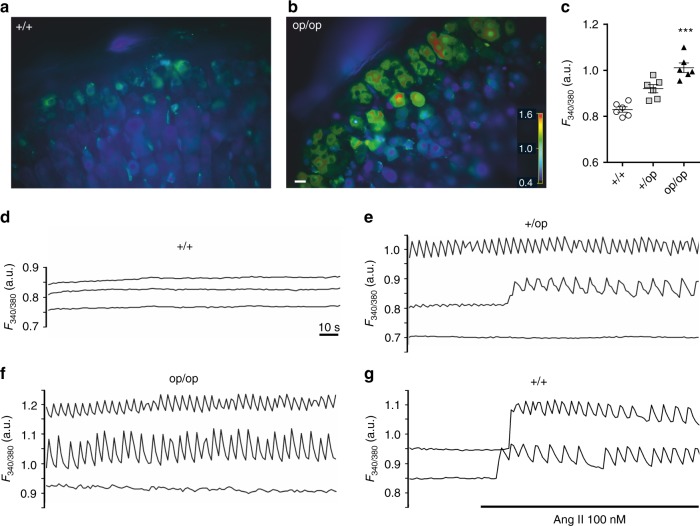
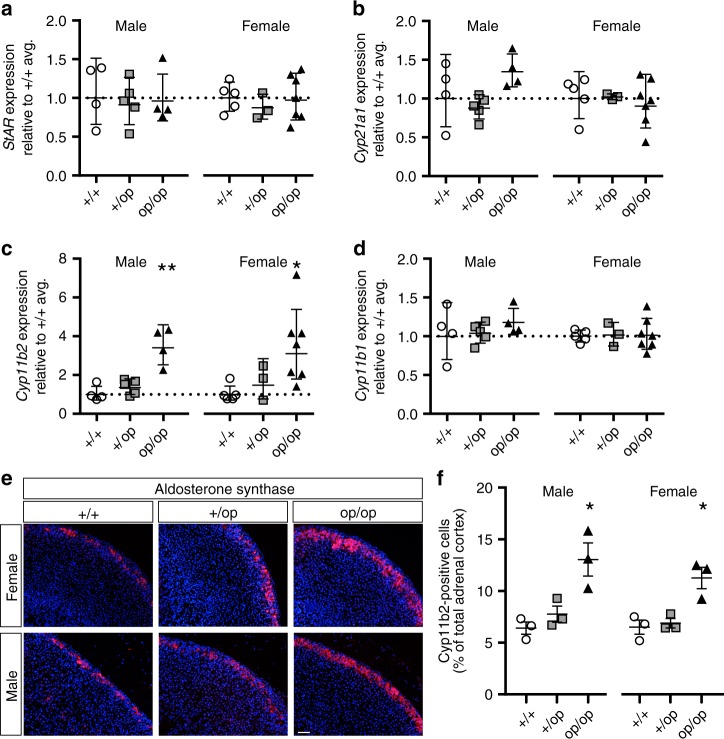
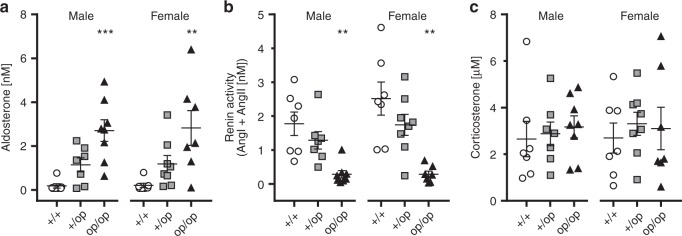
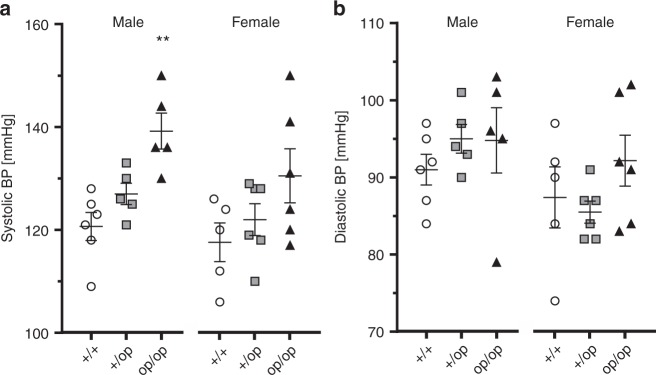
Human primary aldosteronism (PA) can be caused by mutations in several ion channel genes but mouse models replicating this condition are lacking. We now show that almost all known PA-associated CLCN2 mutations markedly increase ClC-2 chloride currents and generate knock-in mice expressing a constitutively open ClC-2 Cl- channel as mouse model for PA. The Clcn2op allele strongly increases the chloride conductance of zona glomerulosa cells, provoking a strong depolarization and increasing cytoplasmic Ca2+ concentration. Clcn2op mice display typical features of human PA, including high serum aldosterone in the presence of low renin activity, marked hypertension and hypokalemia. These symptoms are more pronounced in homozygous Clcn2op/op than in heterozygous Clcn2+/op mice. This difference is attributed to the unexpected finding that only ~50 % of Clcn2+/op zona glomerulosa cells are depolarized. By reproducing essential features of human PA, Clcn2op mice are a valuable model to study the pathological mechanisms underlying this disease.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
A gain-of-function mutation in the CLCN2 chloride channel gene causes primary aldosteronism.Nat Genet. 2018 Mar;50(3):355-361. doi: 10.1038/s41588-018-0053-8. Epub 2018 Feb 5. Nat Genet. 2018. PMID: 29403012
-
Pathogenesis of Familial Hyperaldosteronism Type II: New Concepts Involving Anion Channels.Curr Hypertens Rep. 2019 Apr 4;21(4):31. doi: 10.1007/s11906-019-0934-y. Curr Hypertens Rep. 2019. PMID: 30949771 Review.
-
Adrenal Anion Channels: New Roles in Zona Glomerulosa Physiology and in the Pathophysiology of Primary Aldosteronism.Handb Exp Pharmacol. 2024;283:59-79. doi: 10.1007/164_2023_680. Handb Exp Pharmacol. 2024. PMID: 37495852
-
Elevated aldosterone and blood pressure in a mouse model of familial hyperaldosteronism with ClC-2 mutation.Nat Commun. 2019 Nov 14;10(1):5155. doi: 10.1038/s41467-019-13033-4. Nat Commun. 2019. PMID: 31727896 Free PMC article.
-
New genetic insights in familial hyperaldosteronism.Ann N Y Acad Sci. 2002 Sep;970:77-88. doi: 10.1111/j.1749-6632.2002.tb04414.x. Ann N Y Acad Sci. 2002. PMID: 12381543 Review.
Cited by
-
Intrinsic Adrenal TWIK-Related Acid-Sensitive TASK Channel Dysfunction Produces Spontaneous Calcium Oscillations Sufficient to Drive AngII (Angiotensin II)-Unresponsive Hyperaldosteronism.Hypertension. 2022 Nov;79(11):2552-2564. doi: 10.1161/HYPERTENSIONAHA.122.19557. Epub 2022 Sep 21. Hypertension. 2022. PMID: 36129175 Free PMC article.
-
Ion Channel Function and Electrical Excitability in the Zona Glomerulosa: A Network Perspective on Aldosterone Regulation.Annu Rev Physiol. 2021 Feb 10;83:451-475. doi: 10.1146/annurev-physiol-030220-113038. Epub 2020 Nov 11. Annu Rev Physiol. 2021. PMID: 33176563 Free PMC article. Review.
-
Cellular basis of ClC-2 Cl- channel-related brain and testis pathologies.J Biol Chem. 2021 Jan-Jun;296:100074. doi: 10.1074/jbc.RA120.016031. Epub 2020 Nov 23. J Biol Chem. 2021. PMID: 33187987 Free PMC article.
-
Pathogenesis and treatment of primary aldosteronism.Nat Rev Endocrinol. 2020 Oct;16(10):578-589. doi: 10.1038/s41574-020-0382-4. Epub 2020 Jul 28. Nat Rev Endocrinol. 2020. PMID: 32724183 Review.
-
The Renin-Angiotensin System as a Component of Biotrauma in Acute Respiratory Distress Syndrome.Front Physiol. 2022 Apr 13;12:806062. doi: 10.3389/fphys.2021.806062. eCollection 2021. Front Physiol. 2022. PMID: 35498160 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
