

Bile acids in glucose metabolism and insulin signalling - mechanisms and research needs
- PMID: 31616073
- PMCID: PMC6918475
- DOI: 10.1038/s41574-019-0266-7
Bile acids in glucose metabolism and insulin signalling - mechanisms and research needs
Abstract
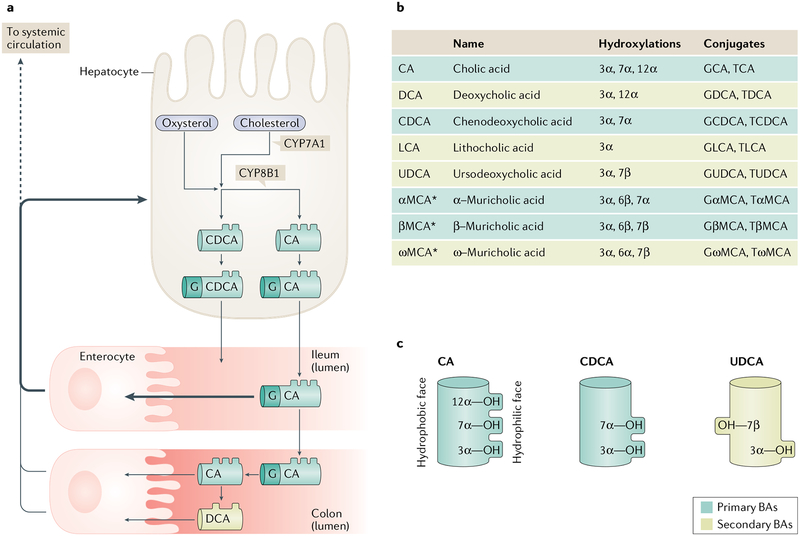
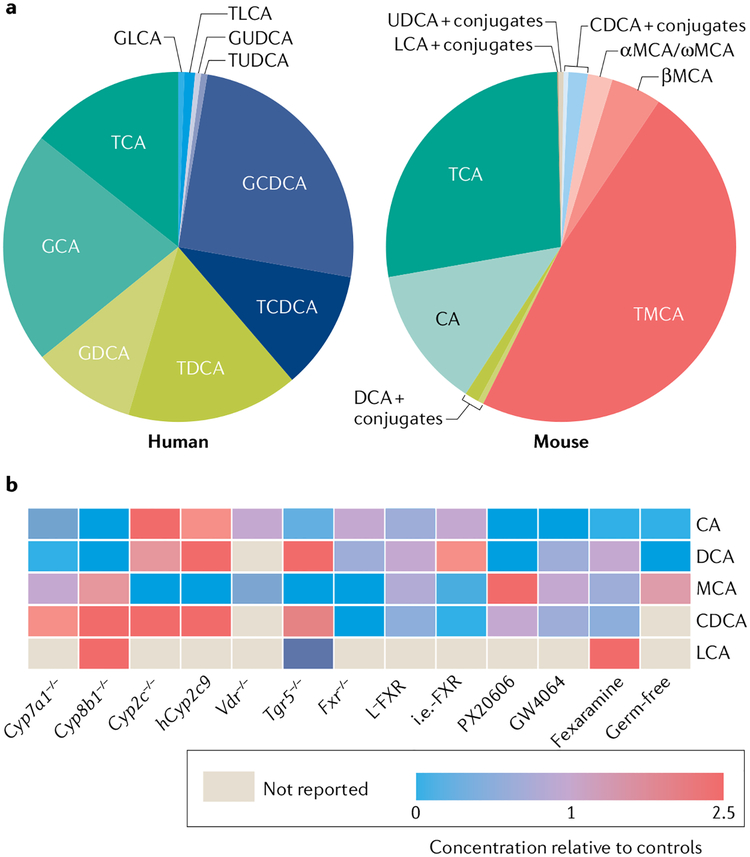
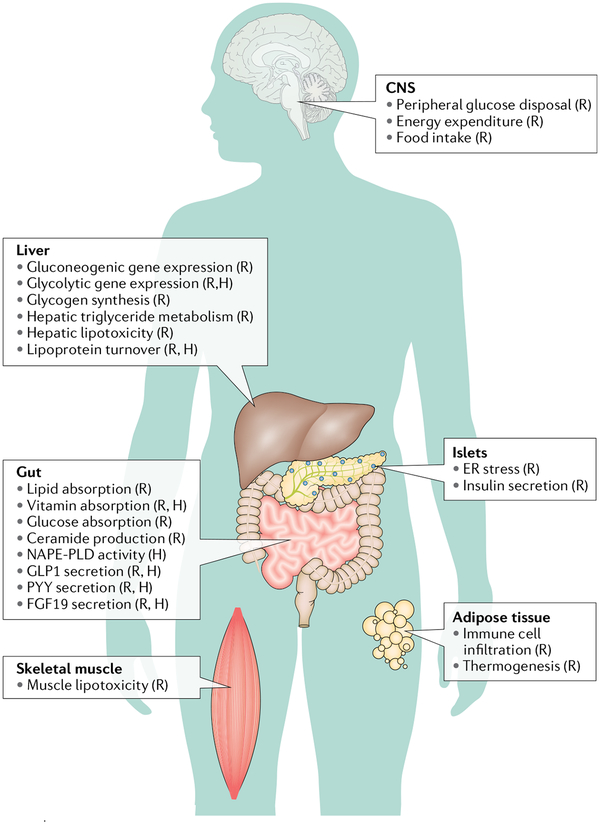
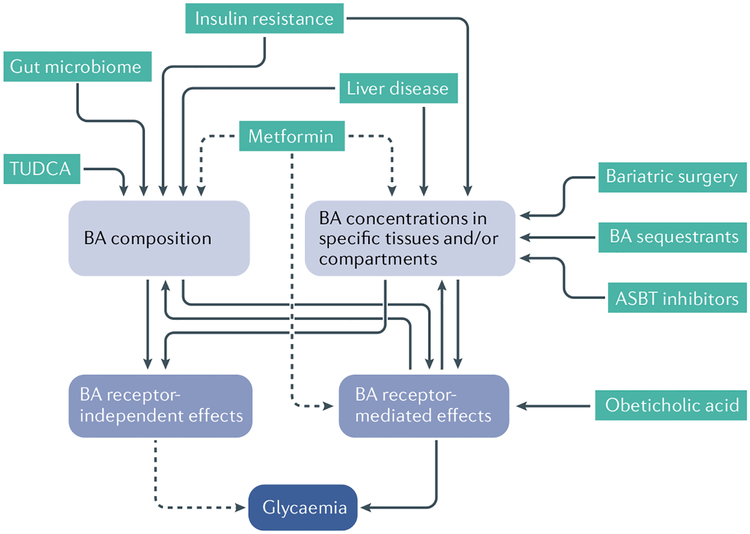
Of all the novel glucoregulatory molecules discovered in the past 20 years, bile acids (BAs) are notable for the fact that they were hiding in plain sight. BAs were well known for their requirement in dietary lipid absorption and biliary cholesterol secretion, due to their micelle-forming properties. However, it was not until 1999 that BAs were discovered to be endogenous ligands for the nuclear receptor FXR. Since that time, BAs have been shown to act through multiple receptors (PXR, VDR, TGR5 and S1PR2), as well as to have receptor-independent mechanisms (membrane dynamics, allosteric modulation of N-acyl phosphatidylethanolamine phospholipase D). We now also have an appreciation of the range of physiological, pathophysiological and therapeutic conditions in which endogenous BAs are altered, raising the possibility that BAs contribute to the effects of these conditions on glycaemia. In this Review, we highlight the mechanisms by which BAs regulate glucose homeostasis and the settings in which endogenous BAs are altered, and provide suggestions for future research.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
References
-
- Hofmann AF The enterohepatic circulation of bile acids in mammals: form and functions. Front. Biosci. Landmark Ed 14, 2584–2598 (2009). - PubMed
-
- Dawson PA in Physiology of the Gastrointestinal Tract 6th edn Ch. 41 (eds Said HM, Ghishan FK, Kaunitz JD, Merchant JL & Wood JD) 931–956 (Academic, 2018).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
