

Different Components of the RNA Interference Machinery Are Required for Conidiation, Ascosporogenesis, Virulence, Deoxynivalenol Production, and Fungal Inhibition by Exogenous Double-Stranded RNA in the Head Blight Pathogen Fusarium graminearum
- PMID: 31616385
- PMCID: PMC6764512
- DOI: 10.3389/fmicb.2019.01662
Different Components of the RNA Interference Machinery Are Required for Conidiation, Ascosporogenesis, Virulence, Deoxynivalenol Production, and Fungal Inhibition by Exogenous Double-Stranded RNA in the Head Blight Pathogen Fusarium graminearum
Abstract
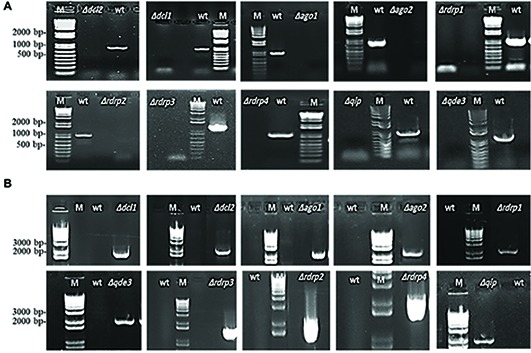
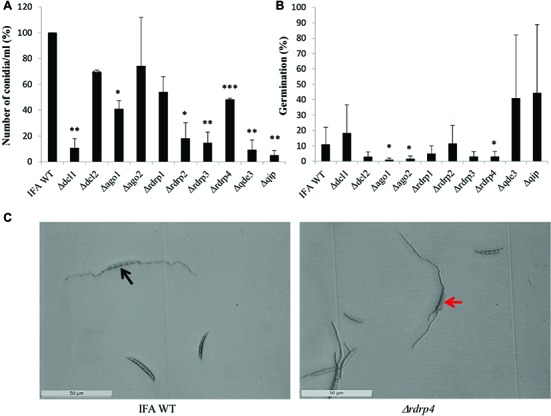
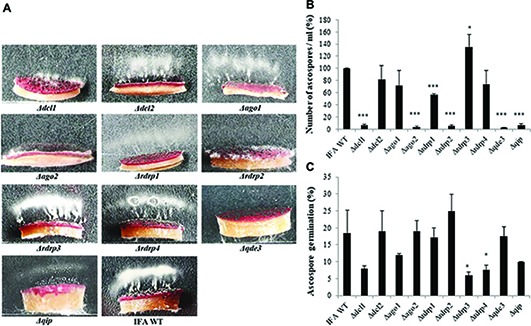
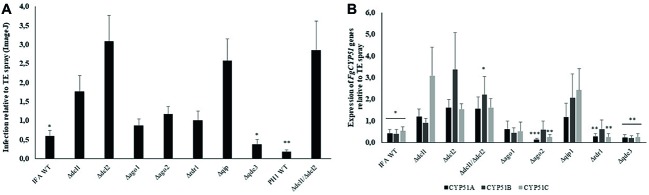
In filamentous fungi, gene silencing through RNA interference (RNAi) shapes many biological processes, including pathogenicity. We explored the requirement of key components of fungal RNAi machineries, including DICER-like 1 and 2 (FgDCL1, FgDCL2), ARGONAUTE 1 and 2 (FgAGO1, FgAGO2), AGO-interacting protein FgQIP (QDE2-interacting protein), RecQ helicase (FgQDE3), and four RNA-dependent RNA polymerases (FgRdRP1, FgRdRP2, FgRdRP3, FgRdRP4), in the ascomycete mycotoxin-producing fungal pathogen Fusarium graminearum (Fg) for sexual and asexual multiplication, pathogenicity, and its sensitivity to double-stranded (ds)RNA. We corroborate and extend earlier findings that conidiation, ascosporogenesis, and Fusarium head blight (FHB) symptom development require an operable RNAi machinery. The involvement of RNAi in conidiation is dependent on environmental conditions as it is detectable only under low light (<2 μmol m-2 s-1). Although both DCLs and AGOs partially share their functions, the sexual ascosporogenesis is mediated primarily by FgDCL1 and FgAGO2, while FgDCL2 and FgAGO1 contribute to asexual conidia formation and germination. FgDCL1 and FgAGO2 also account for pathogenesis as their knockout (KO) results in reduced FHB development. Apart from KO mutants Δdcl2 and Δago1, mutants Δrdrp2, Δrdrp3, Δrdrp4, Δqde3, and Δqip are strongly compromised for conidiation, while KO mutations in all RdPRs, QDE3, and QIP strongly affect ascosporogenesis. Analysis of trichothecenes mycotoxins in wheat kernels showed that the relative amount of deoxynivalenol (DON), calculated as [DON] per amount of fungal genomic DNA was reduced in all spikes infected with RNAi mutants, suggesting the possibility that the fungal RNAi pathways affect Fg's DON production. Moreover, silencing of fungal genes by exogenous target gene-specific double-stranded RNA (dsRNA) (spray-induced gene silencing, SIGS) is dependent on DCLs, AGOs, and QIP, but not on QDE3. Together these data show that in F. graminearum, different key components of the RNAi machinery are crucial in different steps of fungal development and pathogenicity.
Keywords: Argonaute; Fusarium graminearum; double-stranded RNA; small RNA; spray-induced gene silencing; wheat.
Copyright © 2019 Gaffar, Imani, Karlovsky, Koch and Kogel.
Figures

References
LinkOut - more resources
Full Text Sources
Research Materials