HP1 reshapes nucleosome core to promote phase separation of heterochromatin
- PMID: 31618757
- PMCID: PMC7039410
- DOI: 10.1038/s41586-019-1669-2
HP1 reshapes nucleosome core to promote phase separation of heterochromatin
Abstract
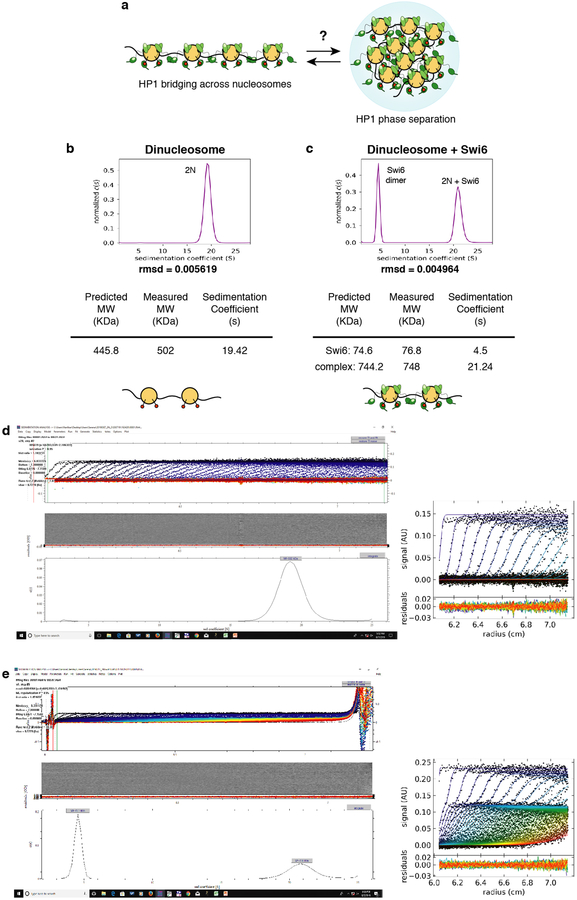
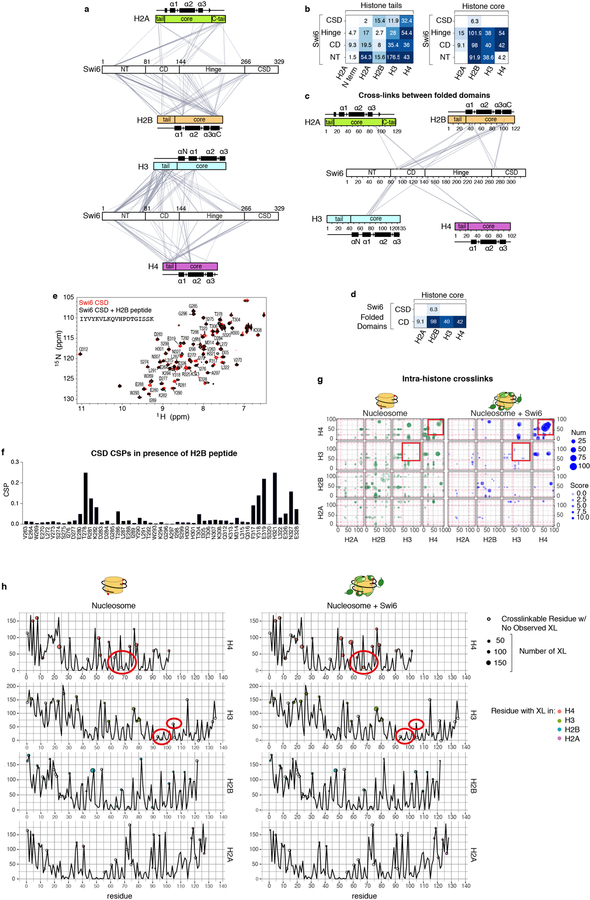
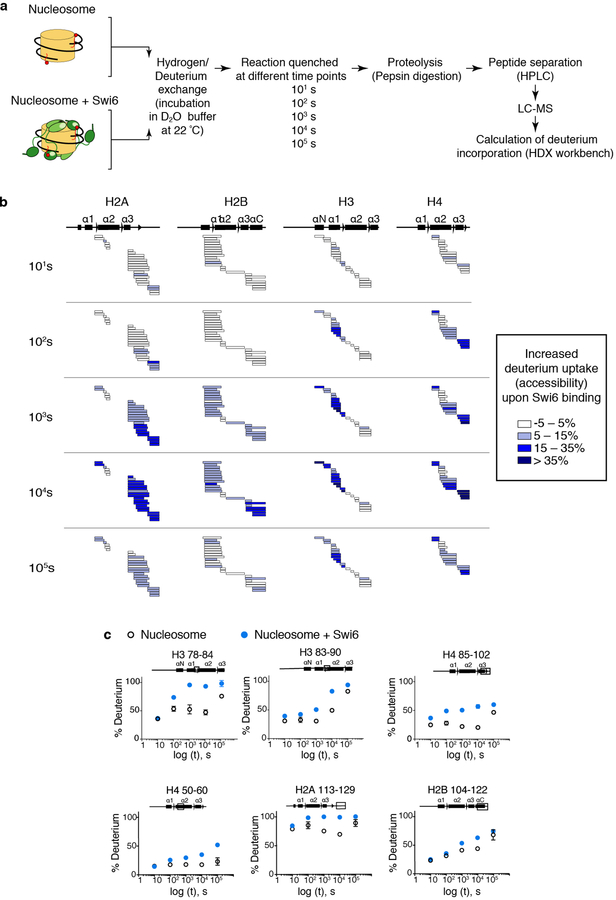
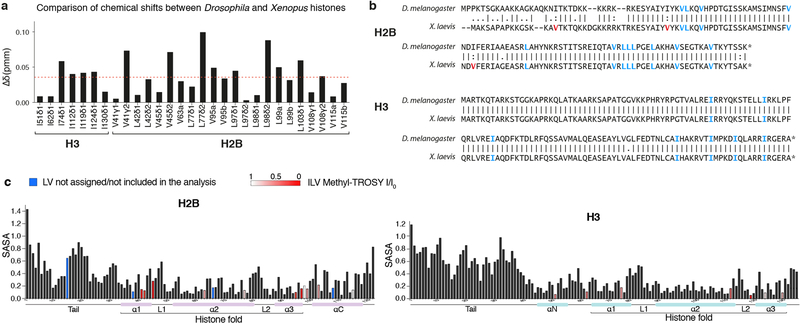
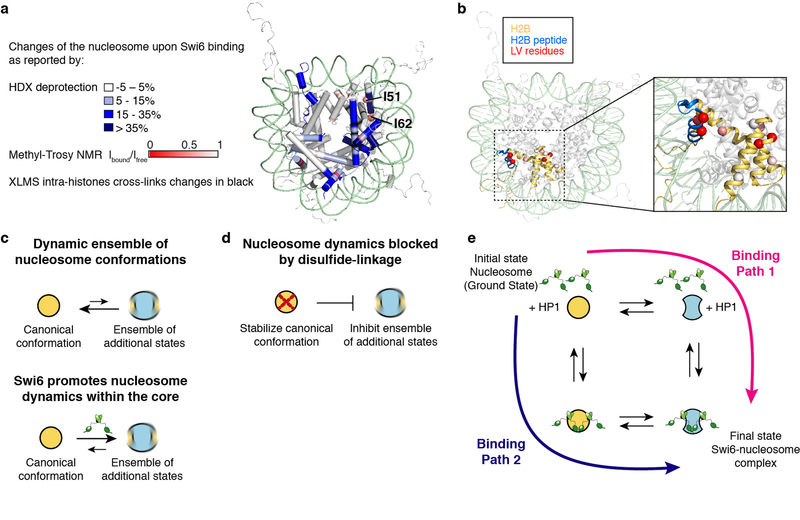
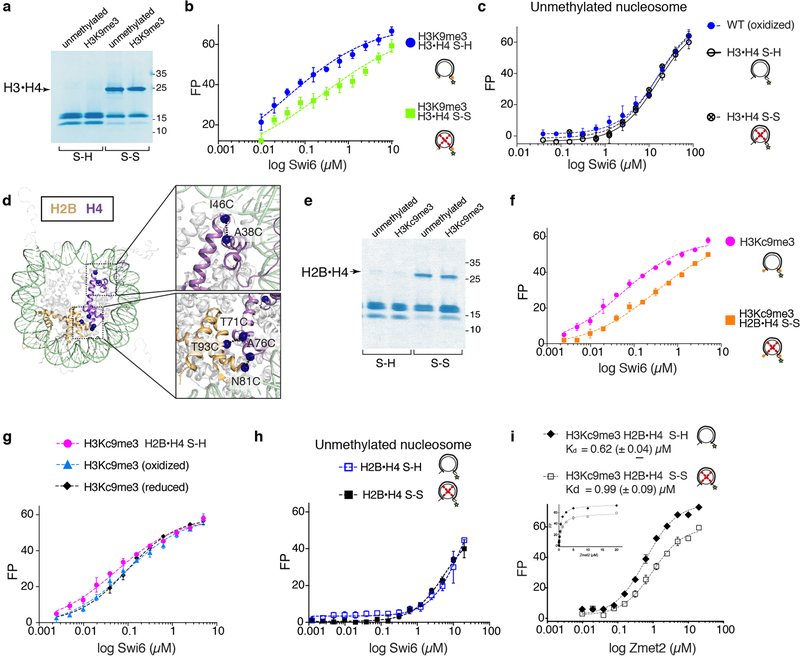
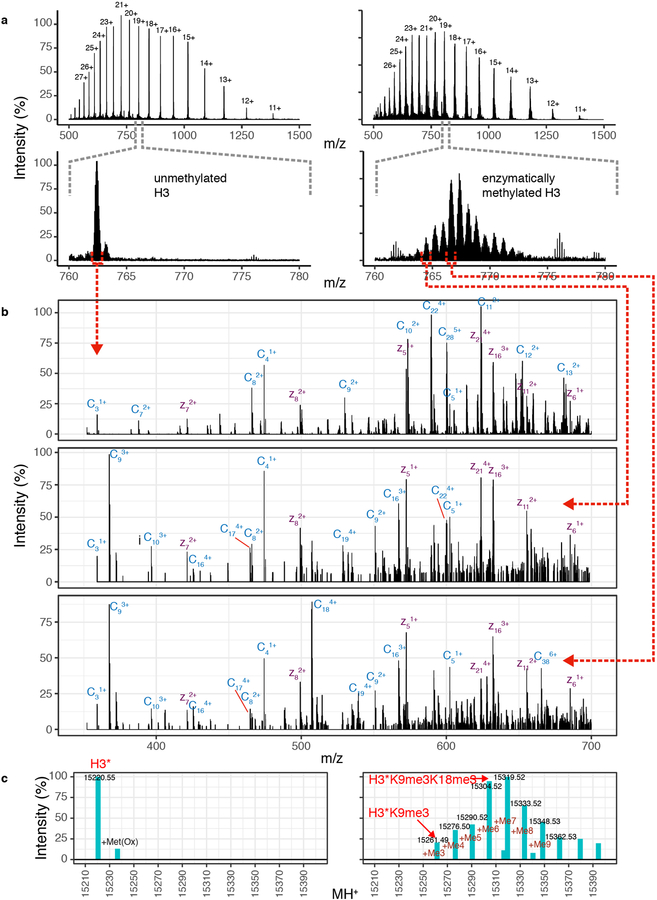
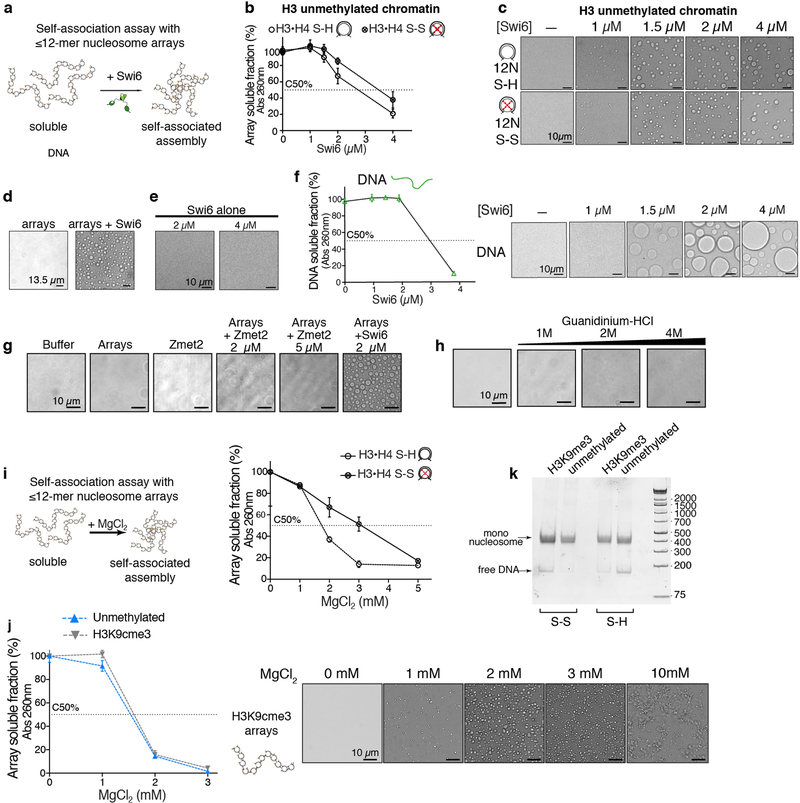
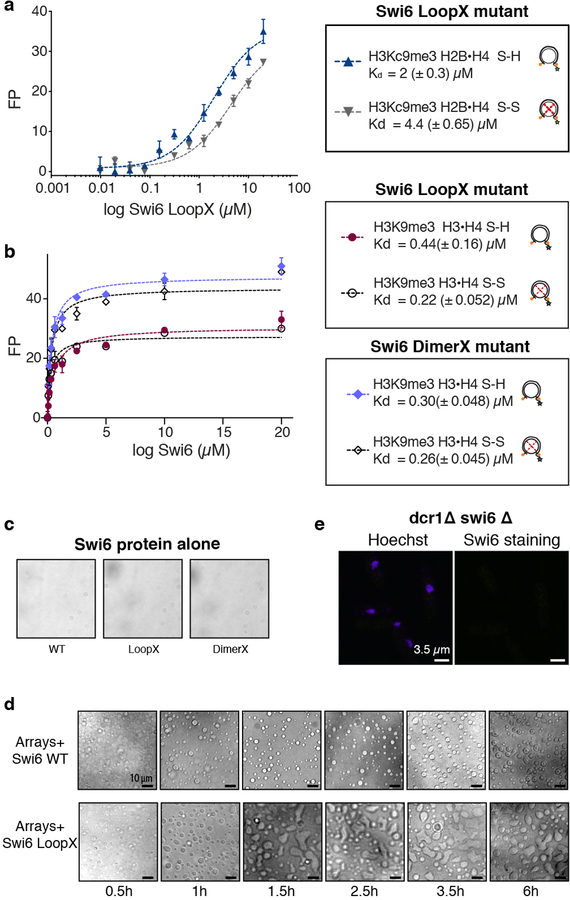
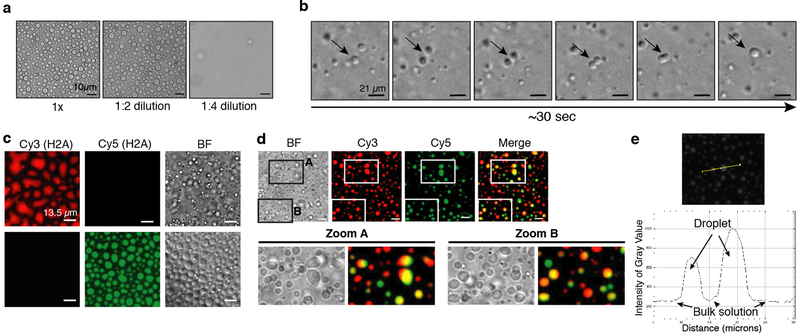
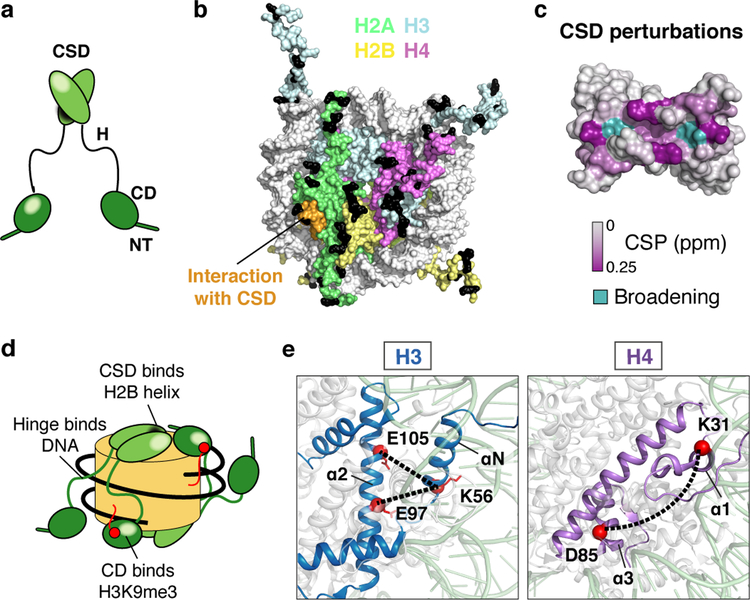
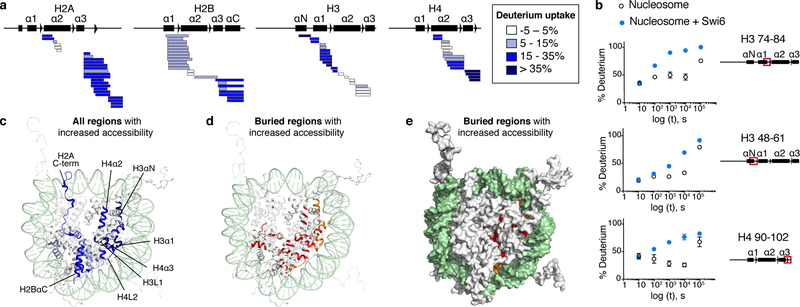
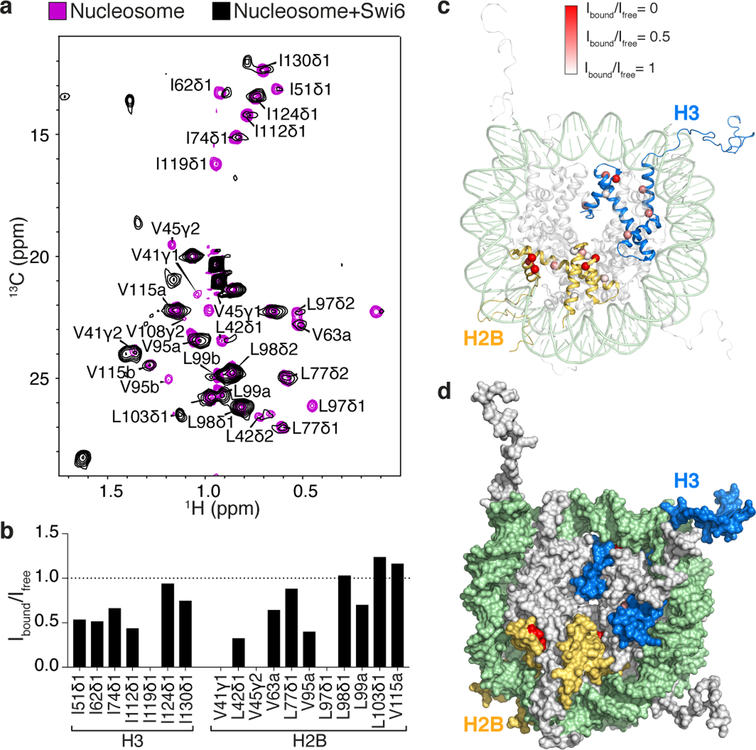
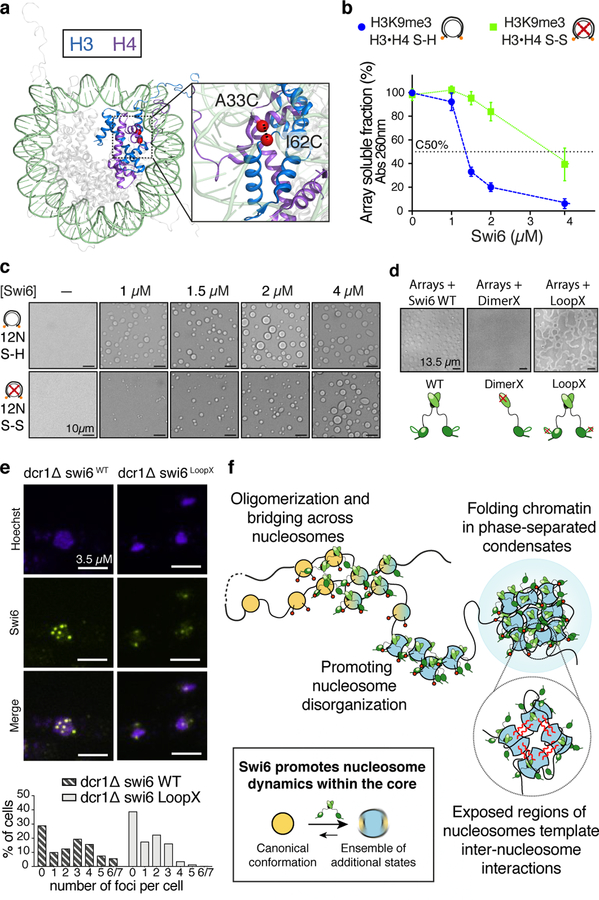
Heterochromatin affects genome function at many levels. It enables heritable gene repression, maintains chromosome integrity and provides mechanical rigidity to the nucleus1,2. These diverse functions are proposed to arise in part from compaction of the underlying chromatin2. A major type of heterochromatin contains at its core the complex formed between HP1 proteins and chromatin that is methylated on histone H3, lysine 9 (H3K9me). HP1 is proposed to use oligomerization to compact chromatin into phase-separated condensates3-6. Yet, how HP1-mediated phase separation relates to chromatin compaction remains unclear. Here we show that chromatin compaction by the Schizosaccharomyces pombe HP1 protein Swi6 results in phase-separated liquid condensates. Unexpectedly, we find that Swi6 substantially increases the accessibility and dynamics of buried histone residues within a nucleosome. Restraining these dynamics impairs compaction of chromatin into liquid droplets by Swi6. Our results indicate that Swi6 couples its oligomerization to the phase separation of chromatin by a counterintuitive mechanism, namely the dynamic exposure of buried nucleosomal regions. We propose that such reshaping of the octamer core by Swi6 increases opportunities for multivalent interactions between nucleosomes, thereby promoting phase separation. This mechanism may more generally drive chromatin organization beyond heterochromatin.
Conflict of interest statement
Competing interests
The authors declare no competing financial interests.
Figures