

Proteostasis collapse is a driver of cell aging and death
- PMID: 31619571
- PMCID: PMC6825304
- DOI: 10.1073/pnas.1906592116
Proteostasis collapse is a driver of cell aging and death
Abstract
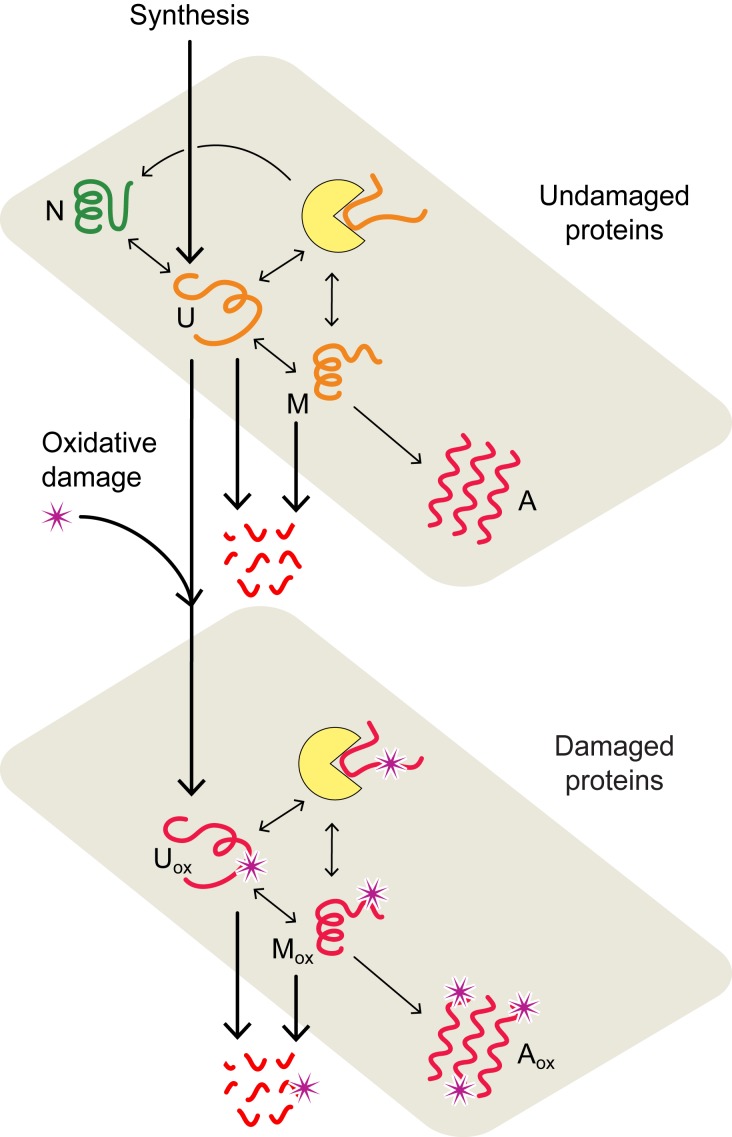
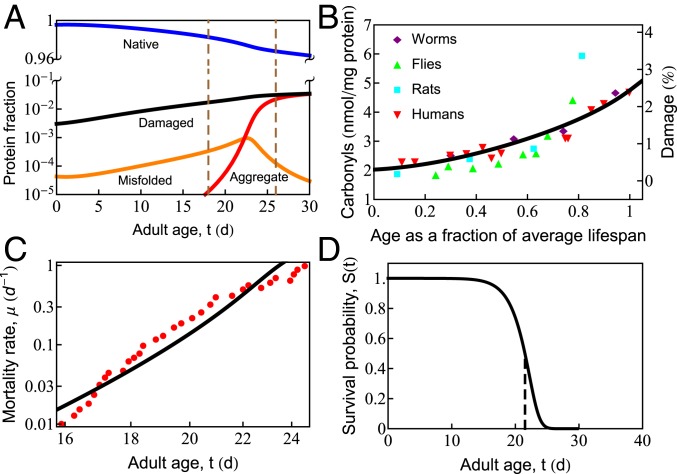
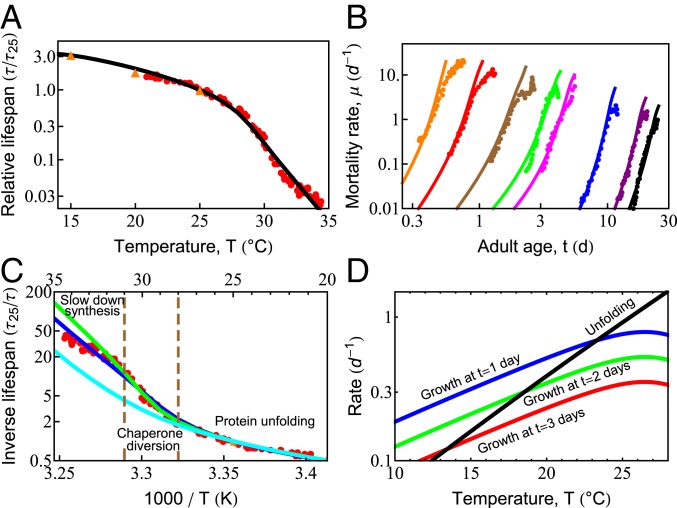
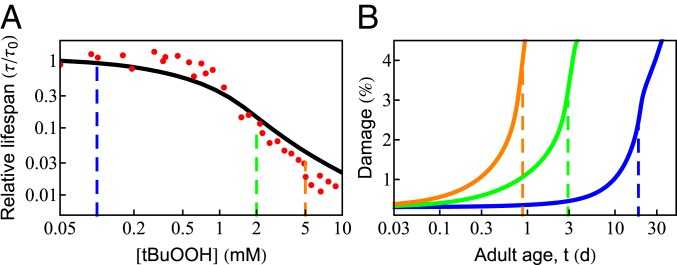
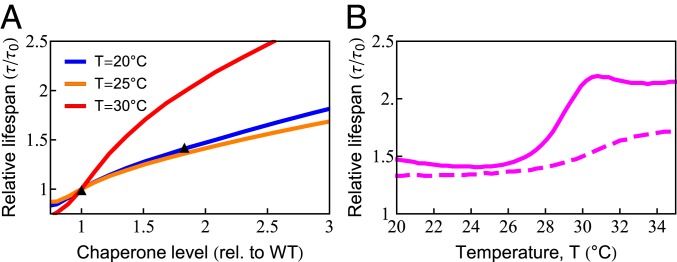
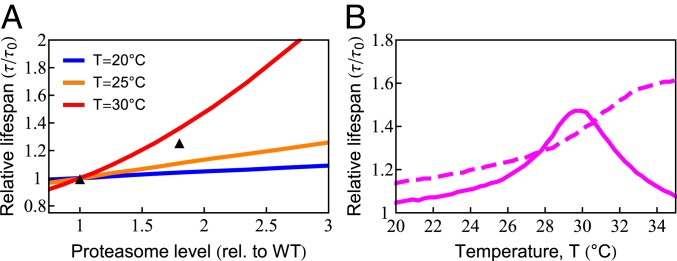
What molecular processes drive cell aging and death? Here, we model how proteostasis-i.e., the folding, chaperoning, and maintenance of protein function-collapses with age from slowed translation and cumulative oxidative damage. Irreparably damaged proteins accumulate with age, increasingly distracting the chaperones from folding the healthy proteins the cell needs. The tipping point to death occurs when replenishing good proteins no longer keeps up with depletion from misfolding, aggregation, and damage. The model agrees with experiments in the worm Caenorhabditis elegans that show the following: Life span shortens nonlinearly with increased temperature or added oxidant concentration, and life span increases in mutants having more chaperones or proteasomes. It predicts observed increases in cellular oxidative damage with age and provides a mechanism for the Gompertz-like rise in mortality observed in humans and other organisms. Overall, the model shows how the instability of proteins sets the rate at which damage accumulates with age and upends a cell's normal proteostasis balance.
Keywords: aging; chaperones; misfolding; oxidative damage; proteostasis.
Copyright © 2019 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Kirkwood T., Understanding the odd science of aging. Cell 120, 437–447 (2005). - PubMed
-
- Kowald A., Kirkwood T., A network theory of ageing: The interactions of defective mitochondria, aberrant proteins, free radicals and scavengers in the ageing process. Mutat. Res. 316, 209–236 (1996). - PubMed
-
- Soti C., Csermely P., Aging and molecular chaperones. Exp. Gerontol. 38, 1037–1040 (2003). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
