

Structural and biochemical characterisation of a novel alginate lyase from Paenibacillus sp. str. FPU-7
- PMID: 31619701
- PMCID: PMC6796002
- DOI: 10.1038/s41598-019-51006-1
Structural and biochemical characterisation of a novel alginate lyase from Paenibacillus sp. str. FPU-7
Abstract
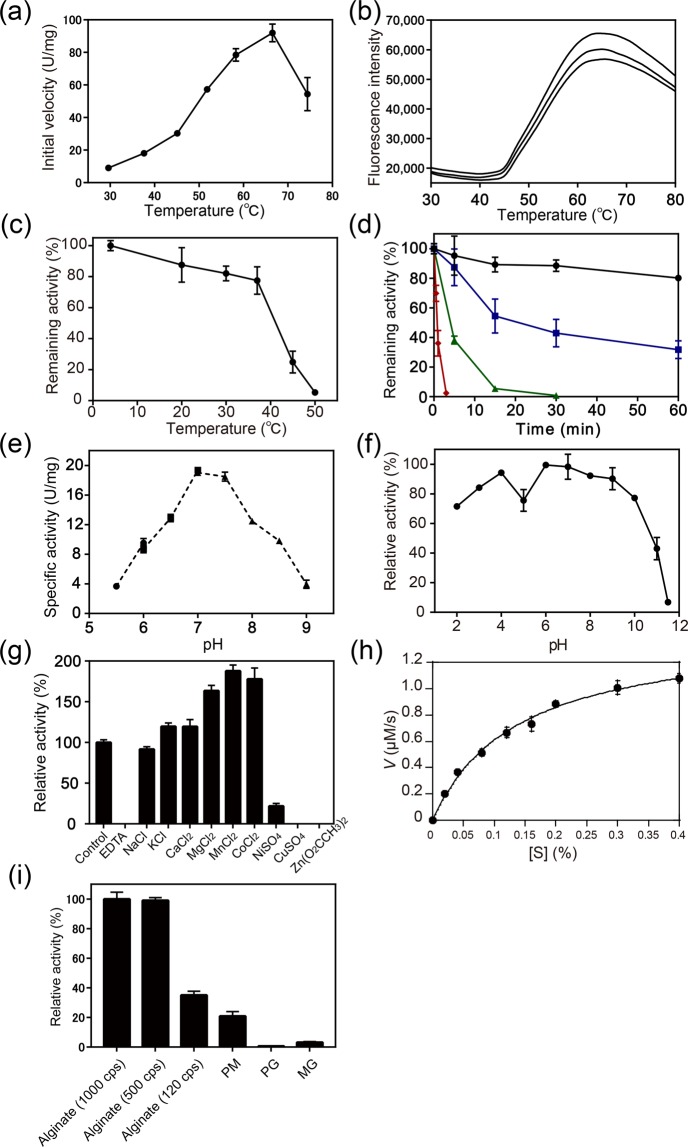
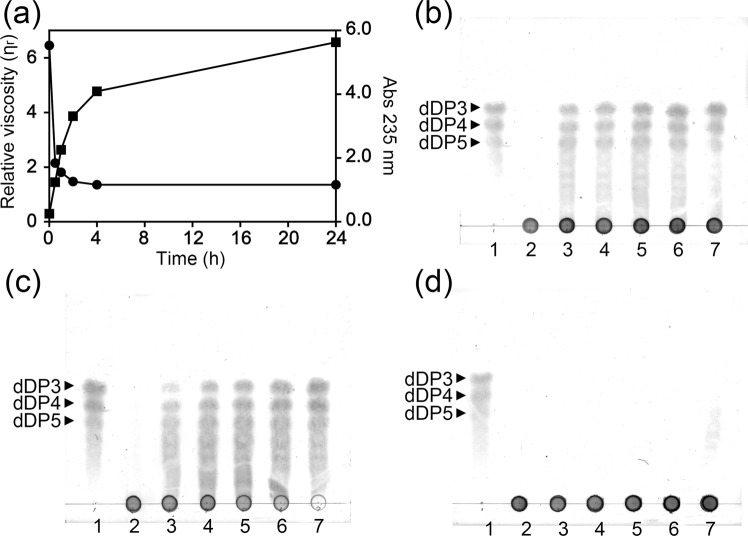
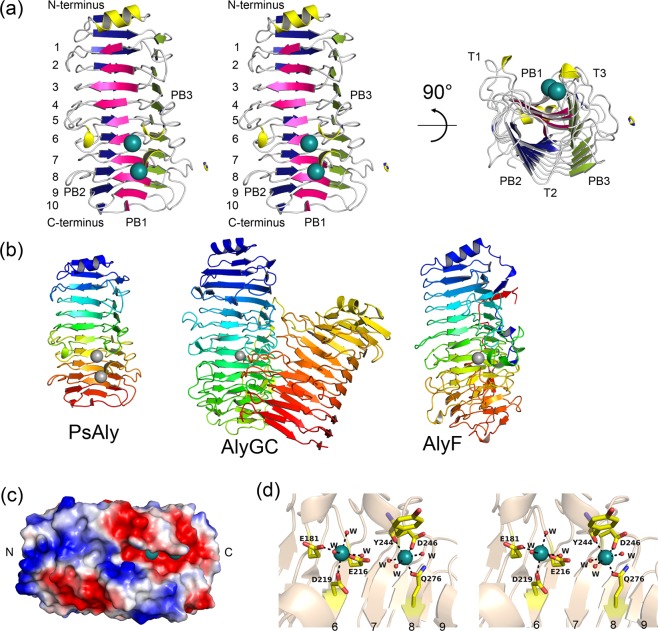
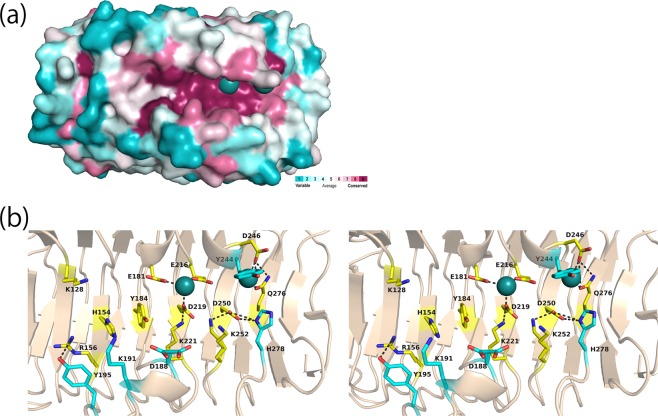
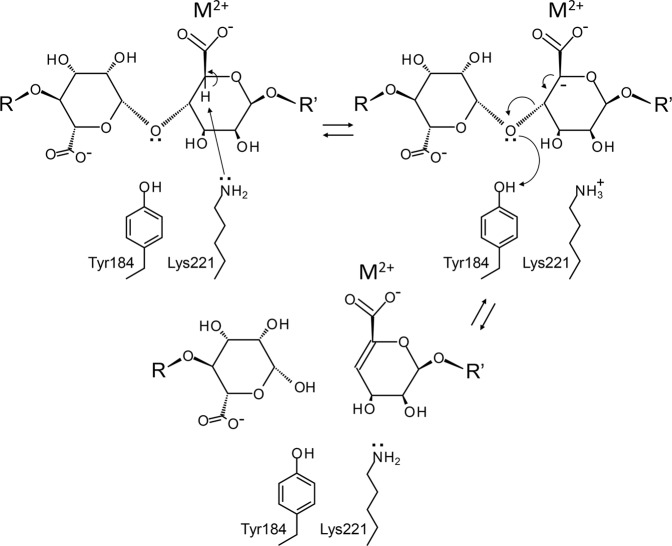
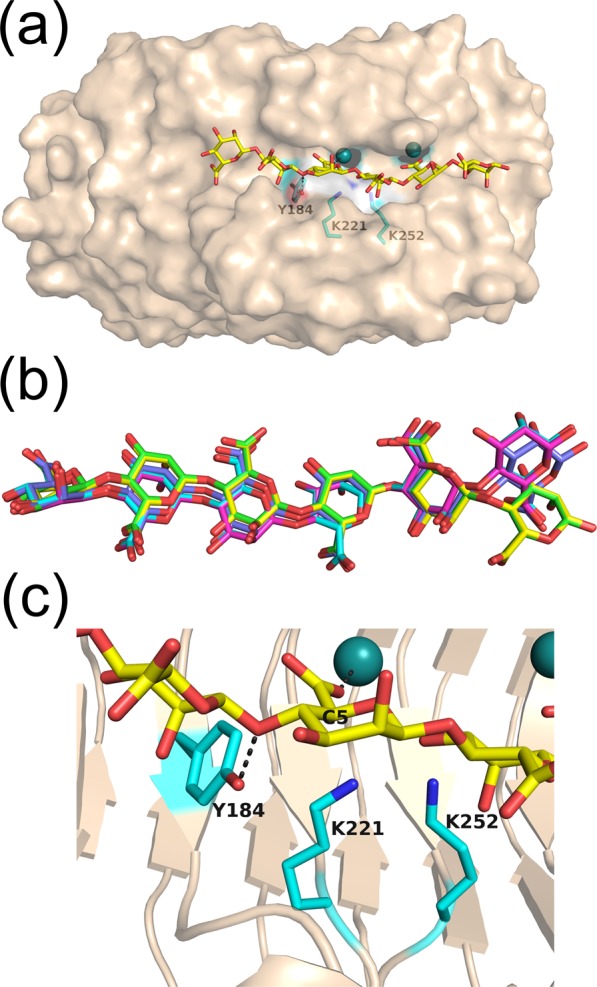
A novel alginate lyase, PsAly, with a molecular mass of 33 kDa and whose amino acid sequence shares no significant similarity to other known proteins, was biochemically and structurally characterised from Paenibacillus sp. str. FPU-7. The maximum PsAly activity was obtained at 65 °C, with an optimum pH of pH 7-7.5. The activity was enhanced by divalent cations, such as Mg2+, Mn2+, or Co2+, and inhibited by a metal chelator, ethylenediaminetetraacetic acid. The reaction products indicated that PsAly is an endolytic enzyme with a preference for polymannuronate. Herein, we report a detailed crystal structure of PsAly at a resolution of 0.89 Å, which possesses a β-helix fold that creates a long cleft. The catalytic site was different from that of other polysaccharide lyases. Site-directed mutational analysis of conserved residues predicted Tyr184 and Lys221 as catalytic residues, abstracting from the C5 proton and providing a proton to the glycoside bond, respectively. One cation was found to bind to the bottom of the cleft and neutralise the carboxy group of the substrate, decreasing the pKa of the C5 proton to promote catalysis. Our study provides an insight into the structural basis for the catalysis of alginate lyases and β-helix polysaccharide lyases.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
