

Filament-like Assemblies of Intracellular Nucleic Acid Sensors: Commonalities and Differences
- PMID: 31626748
- PMCID: PMC6880955
- DOI: 10.1016/j.molcel.2019.09.023
Filament-like Assemblies of Intracellular Nucleic Acid Sensors: Commonalities and Differences
Abstract
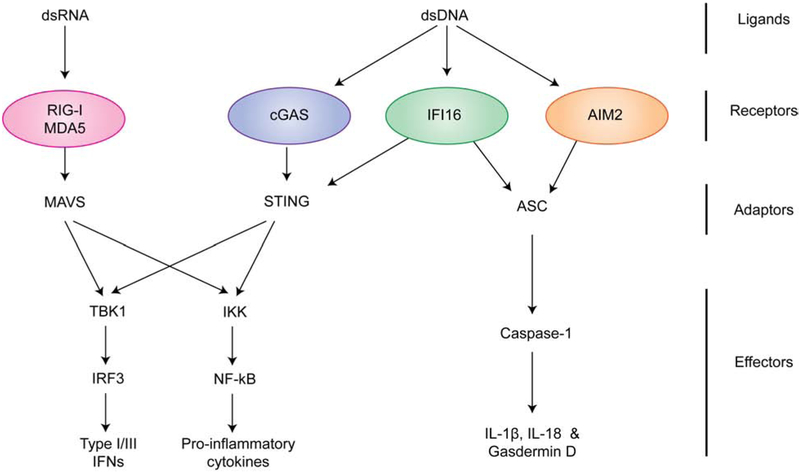
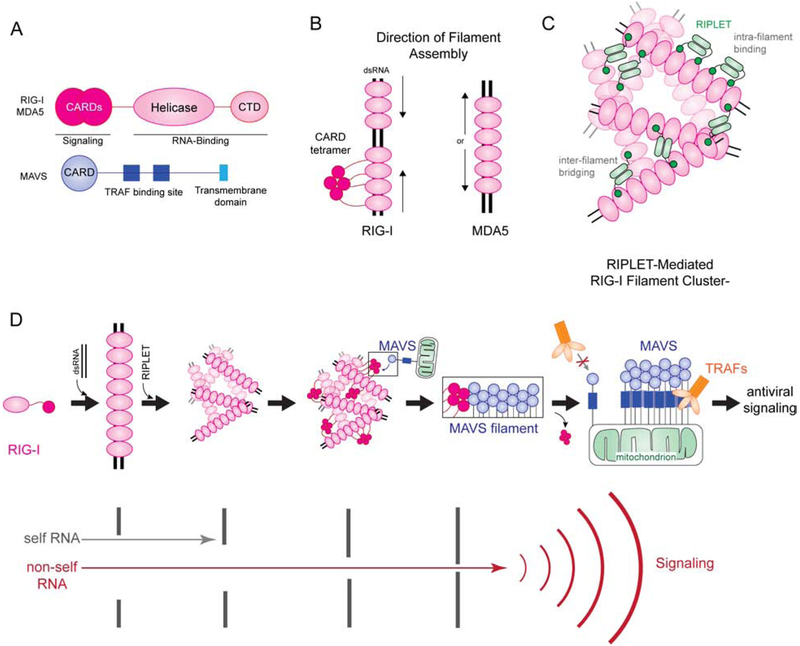
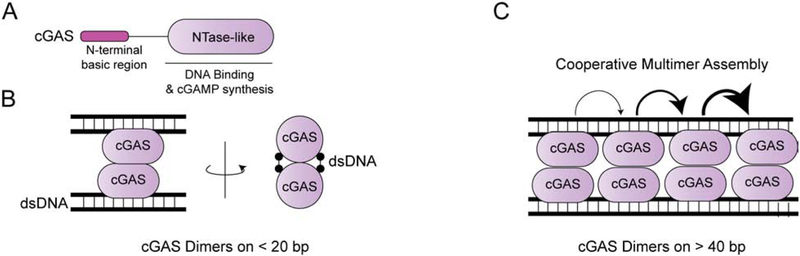
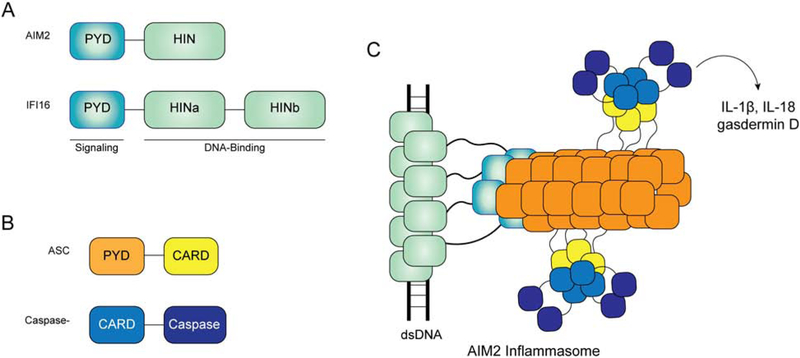
Self versus non-self discrimination by innate immune sensors is critical for mounting effective immune responses against pathogens while avoiding harmful auto-inflammatory reactions against the host. Foreign DNA and RNA sensors must discriminate between self versus non-self nucleic acids, despite their shared building blocks and similar physicochemical properties. Recent structural and biochemical studies suggest that multiple steps of filament-like assembly are required for the functions of several nucleic acid sensors. Here, we discuss ligand discrimination and oligomerization of RIG-I-like receptors, AIM2-like receptors, and cGAS. We discuss how filament-like assembly allows for robust and accurate discrimination of self versus non-self nucleic acids and how these assemblies enable sensing of multiple distinct features in foreign nucleic acids, including structure, length, and modifications. We also discuss how individual receptors differ in their assembly and disassembly mechanisms and how these differences contribute to the diversity in nucleic acid specificity and pathogen detection strategies.
Copyright © 2019 Elsevier Inc. All rights reserved.
Figures
References
-
- Ablasser A, and Chen ZJ (2019). cGAS in action: Expanding roles in immunity and inflammation. Science 363. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
