

Reformulating Pro-Oxidant Microglia in Neurodegeneration
- PMID: 31627485
- PMCID: PMC6832973
- DOI: 10.3390/jcm8101719
Reformulating Pro-Oxidant Microglia in Neurodegeneration
Abstract
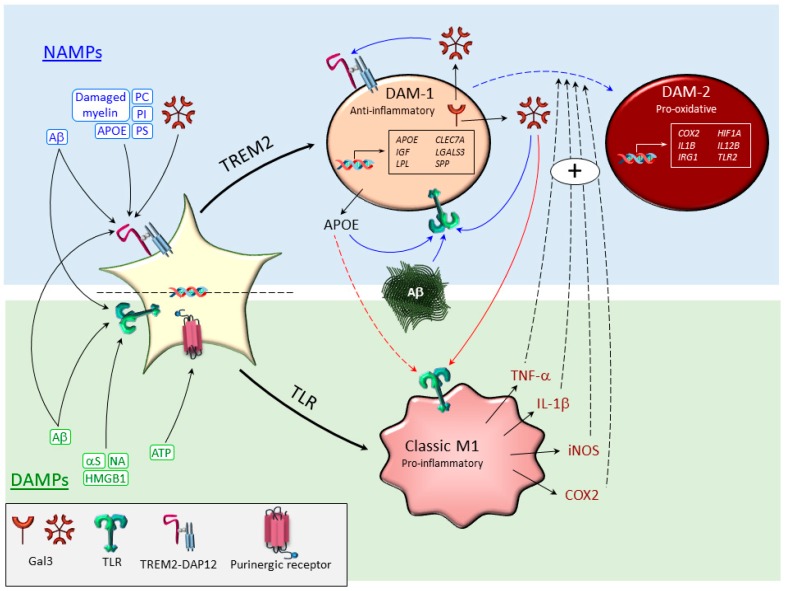
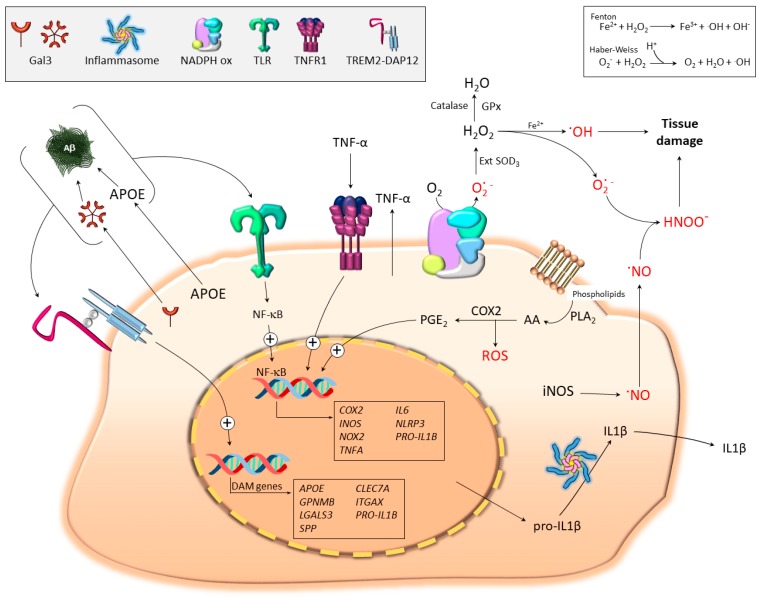
In neurodegenerative diseases, microglia-mediated neuroinflammation and oxidative stress are central events. Recent genome-wide transcriptomic analyses of microglial cells under different disease conditions have uncovered a new subpopulation named disease-associated microglia (DAM). These studies have challenged the classical view of the microglia polarization state's proinflammatory M1 (classical activation) and immunosuppressive M2 (alternative activation). Molecular signatures of DAM and proinflammatory microglia (highly pro-oxidant) have shown clear differences, yet a partial overlapping gene profile is evident between both phenotypes. The switch activation of homeostatic microglia into reactive microglia relies on the selective activation of key surface receptors involved in the maintenance of brain homeostasis (a.k.a. pattern recognition receptors, PRRs). Two relevant PRRs are toll-like receptors (TLRs) and triggering receptors expressed on myeloid cells-2 (TREM2), whose selective activation is believed to generate either a proinflammatory or a DAM phenotype, respectively. However, the recent identification of endogenous disease-related ligands, which bind to and activate both TLRs and TREM2, anticipates the existence of rather complex microglia responses. Examples of potential endogenous dual ligands include amyloid β, galectin-3, and apolipoprotein E. These pleiotropic ligands induce a microglia polarization that is more complicated than initially expected, suggesting the possibility that different microglia subtypes may coexist. This review highlights the main microglia polarization states under disease conditions and their leading role orchestrating oxidative stress.
Keywords: RNS; ROS; disease-associated microglia (DAM); inflammation; microglia; neurodegeneration; oxidative stress.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Del Rio-Hortega P. El tercer elemento de los centros nerviosos. Bol. Soc. Esp. Biol. 1919;9:69–120.
-
- Del Rio-Hortega P. The microglia. Lancet. 1939;1:1023–1026. doi: 10.1016/S0140-6736(00)60571-8. - DOI
