

Three Rostromedial Tegmental Afferents Drive Triply Dissociable Aspects of Punishment Learning and Aversive Valence Encoding
- PMID: 31627985
- PMCID: PMC6989096
- DOI: 10.1016/j.neuron.2019.08.040
Three Rostromedial Tegmental Afferents Drive Triply Dissociable Aspects of Punishment Learning and Aversive Valence Encoding
Abstract
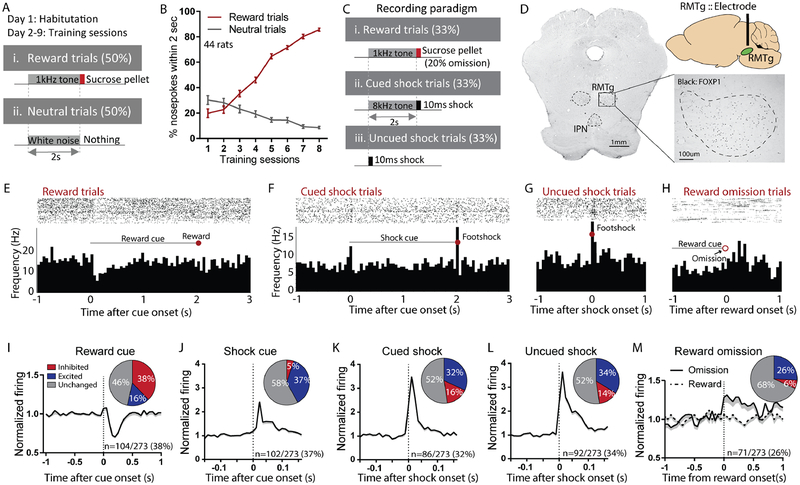
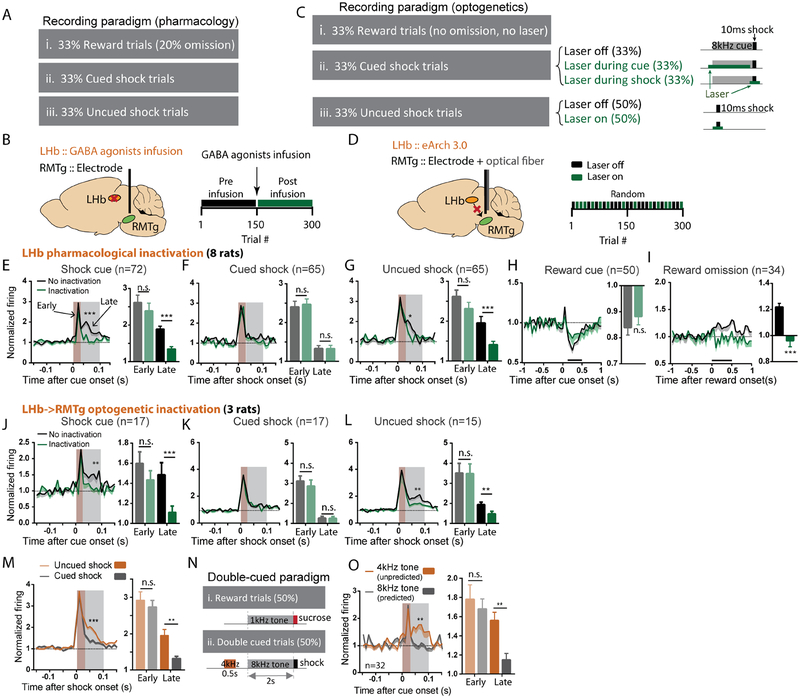
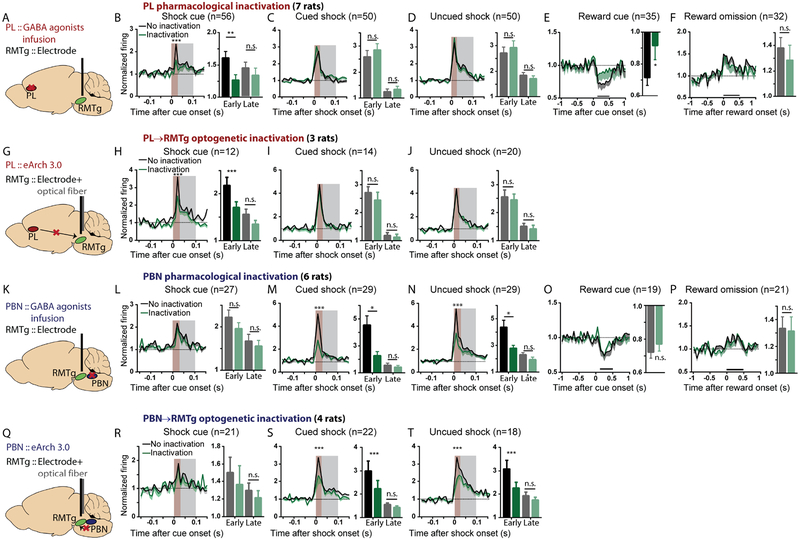
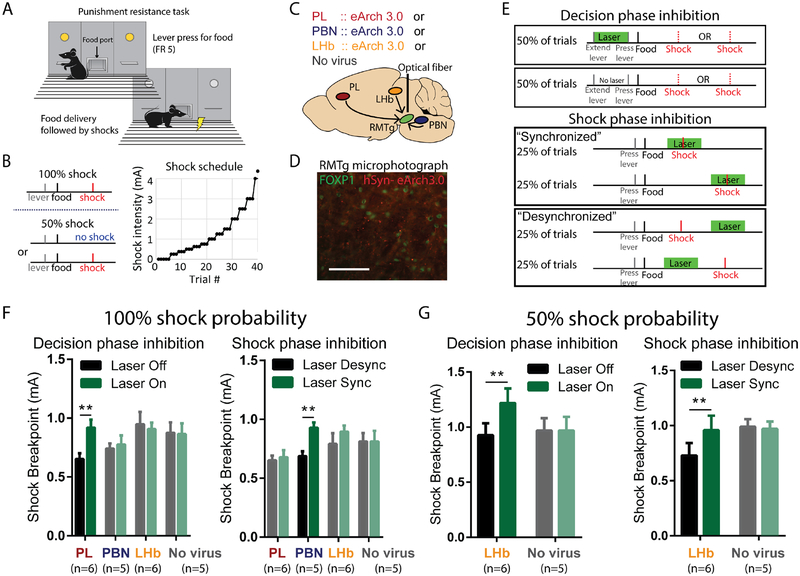
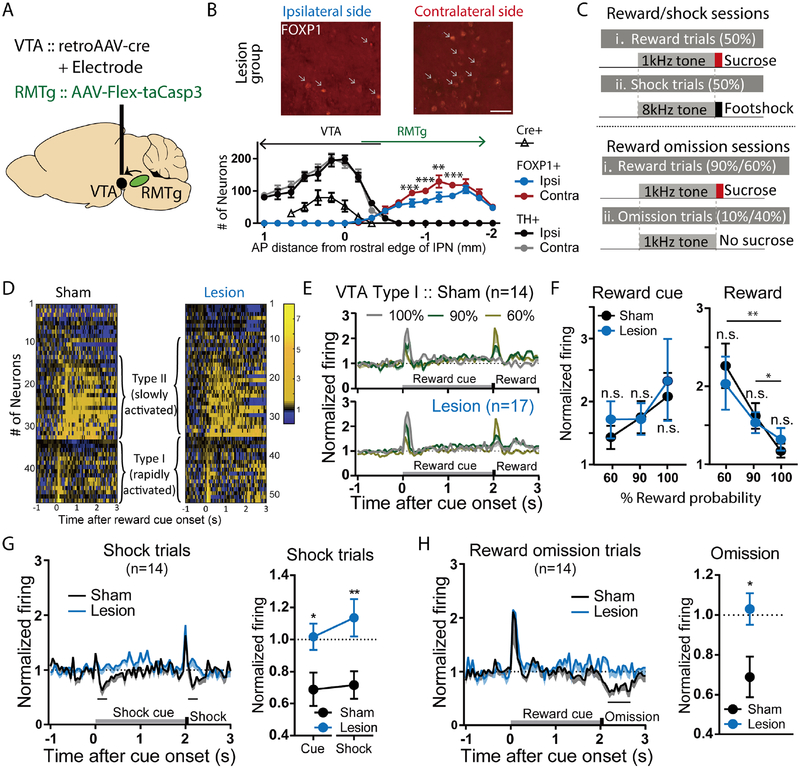
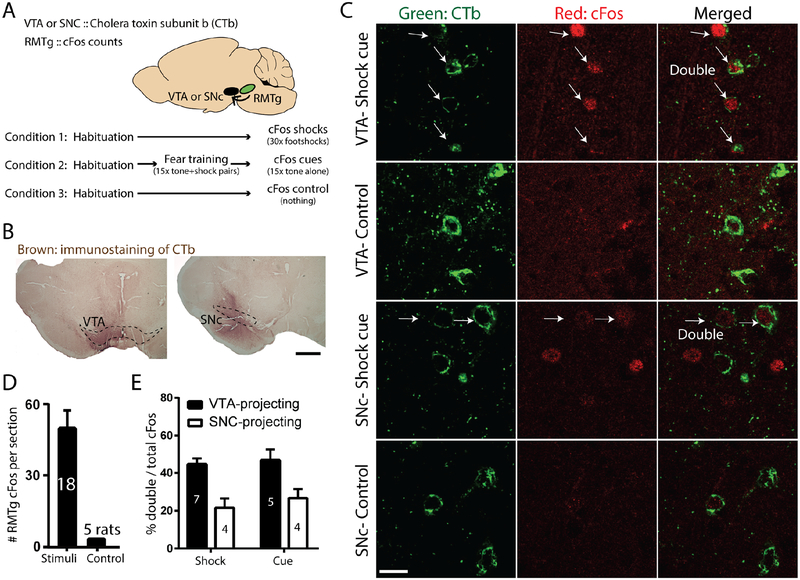
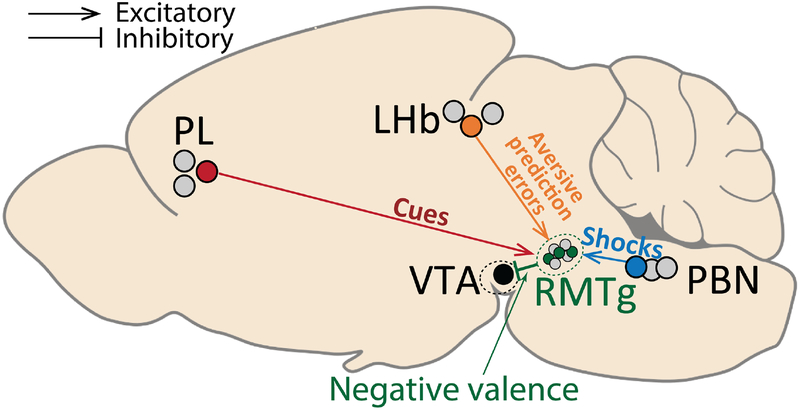
Persistence of reward seeking despite punishment or other negative consequences is a defining feature of mania and addiction, and numerous brain regions have been implicated in such punishment learning, but in disparate ways that are difficult to reconcile. We now show that the ability of an aversive punisher to inhibit reward seeking depends on coordinated activity of three distinct afferents to the rostromedial tegmental nucleus (RMTg) arising from cortex, brainstem, and habenula that drive triply dissociable RMTg responses to aversive cues, outcomes, and prediction errors, respectively. These three pathways drive correspondingly dissociable aspects of punishment learning. The RMTg in turn drives negative, but not positive, valence encoding patterns in the ventral tegmental area (VTA). Hence, punishment learning involves pathways and functions that are highly distinct, yet tightly coordinated.
Keywords: RMTg; VTA; aversion; dopamine; habenula; learning; parabrachial; prefrontal; punishment; valence.
Copyright © 2019 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests
No conflicts in interests
Figures
References
-
- American Psychiatric Association; (2013). Diagnostic and Statistical Manual of Mental Disorders, 5th edn (Washington, DC: ).
-
- Baker PM, Raynor SA, Francis NT, and Mizumori SJ (2017). Lateral habenula integration of proactive and retroactive information mediates behavioral flexibility. Neuroscience 345, 89–98. - PubMed
-
- Bourdy R, Sanchez-Catalan MJ, Kaufling J, Balcita-Pedicino JJ, Freund-Mercier MJ, Veinante P, Sesack SR, Georges F, and Barrot M (2014). Control of the nigrostriatal dopamine neuron activity and motor function by the tail of the ventral tegmental area. Neuropsychopharmacology 39, 2788–2798. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
