

A Promiscuous CYP706A3 Reduces Terpene Volatile Emission from Arabidopsis Flowers, Affecting Florivores and the Floral Microbiome
- PMID: 31628167
- PMCID: PMC6925022
- DOI: 10.1105/tpc.19.00320
A Promiscuous CYP706A3 Reduces Terpene Volatile Emission from Arabidopsis Flowers, Affecting Florivores and the Floral Microbiome
Abstract
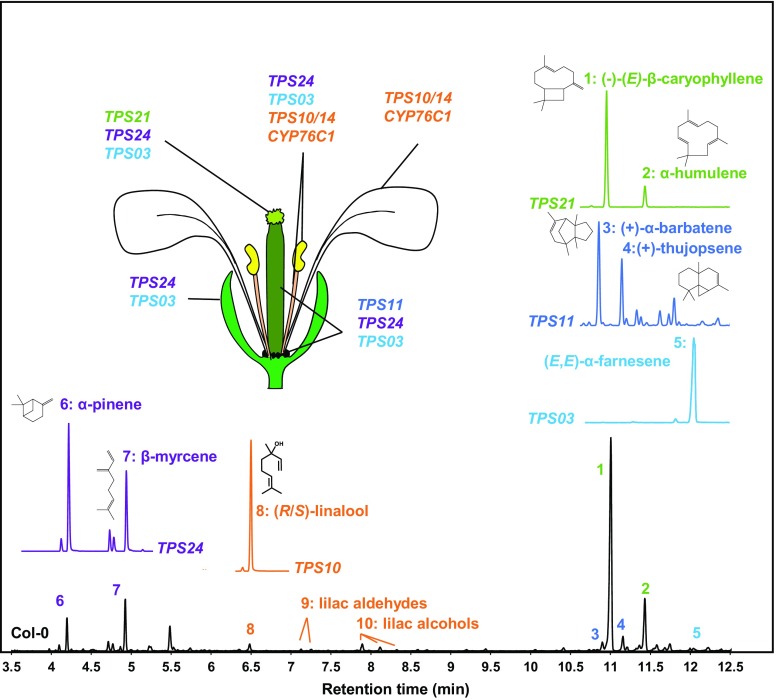
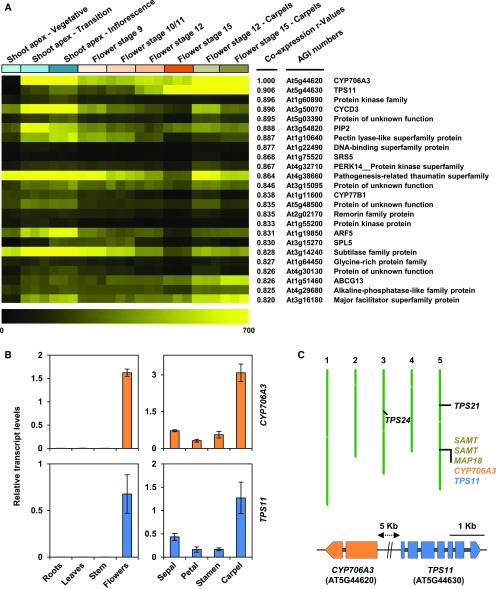
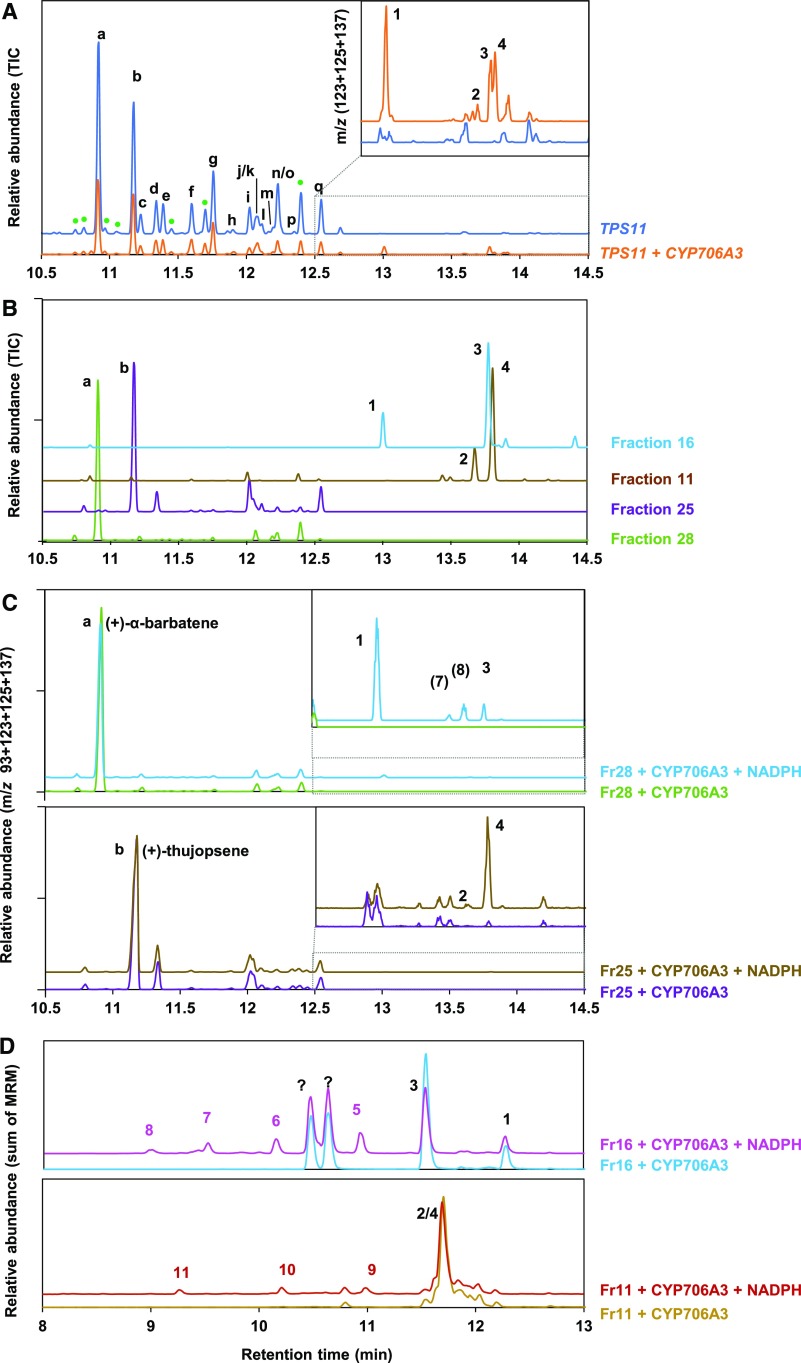
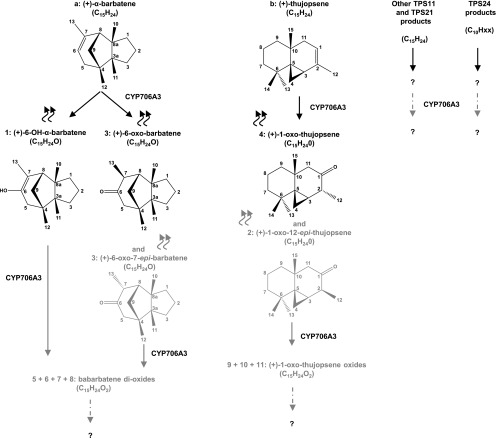
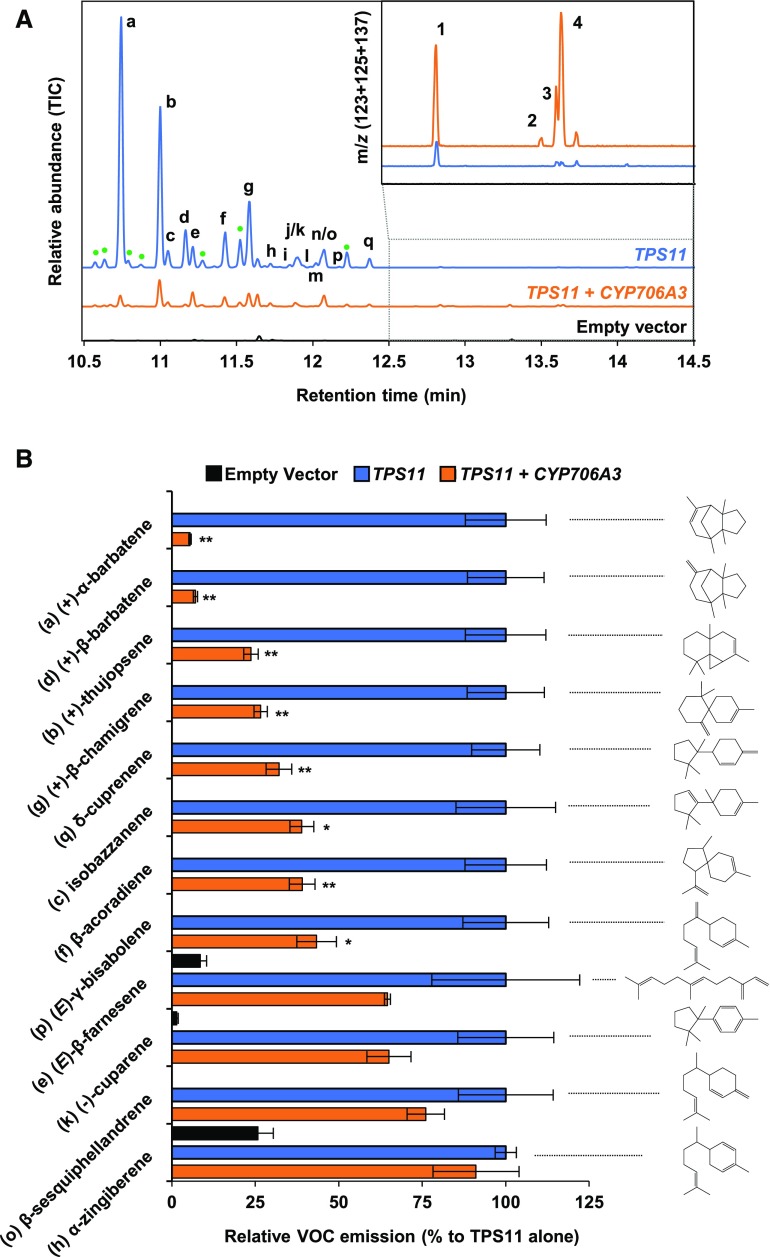
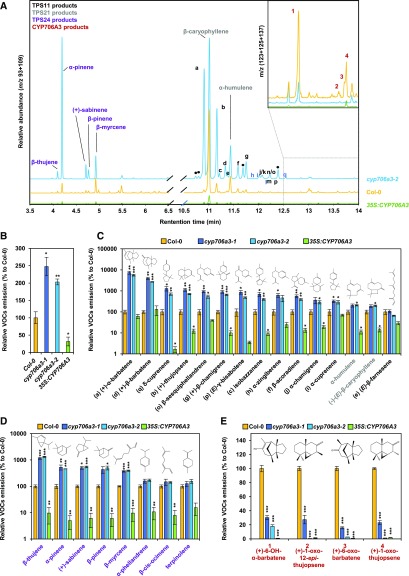
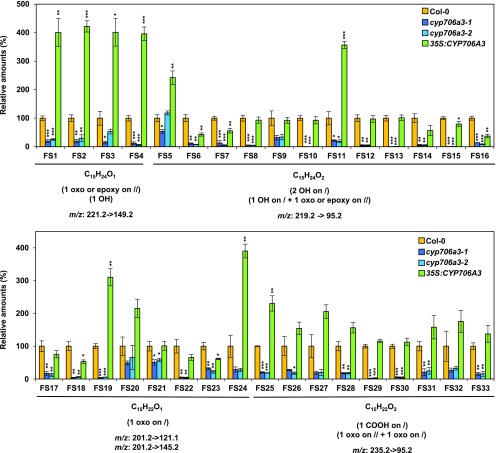
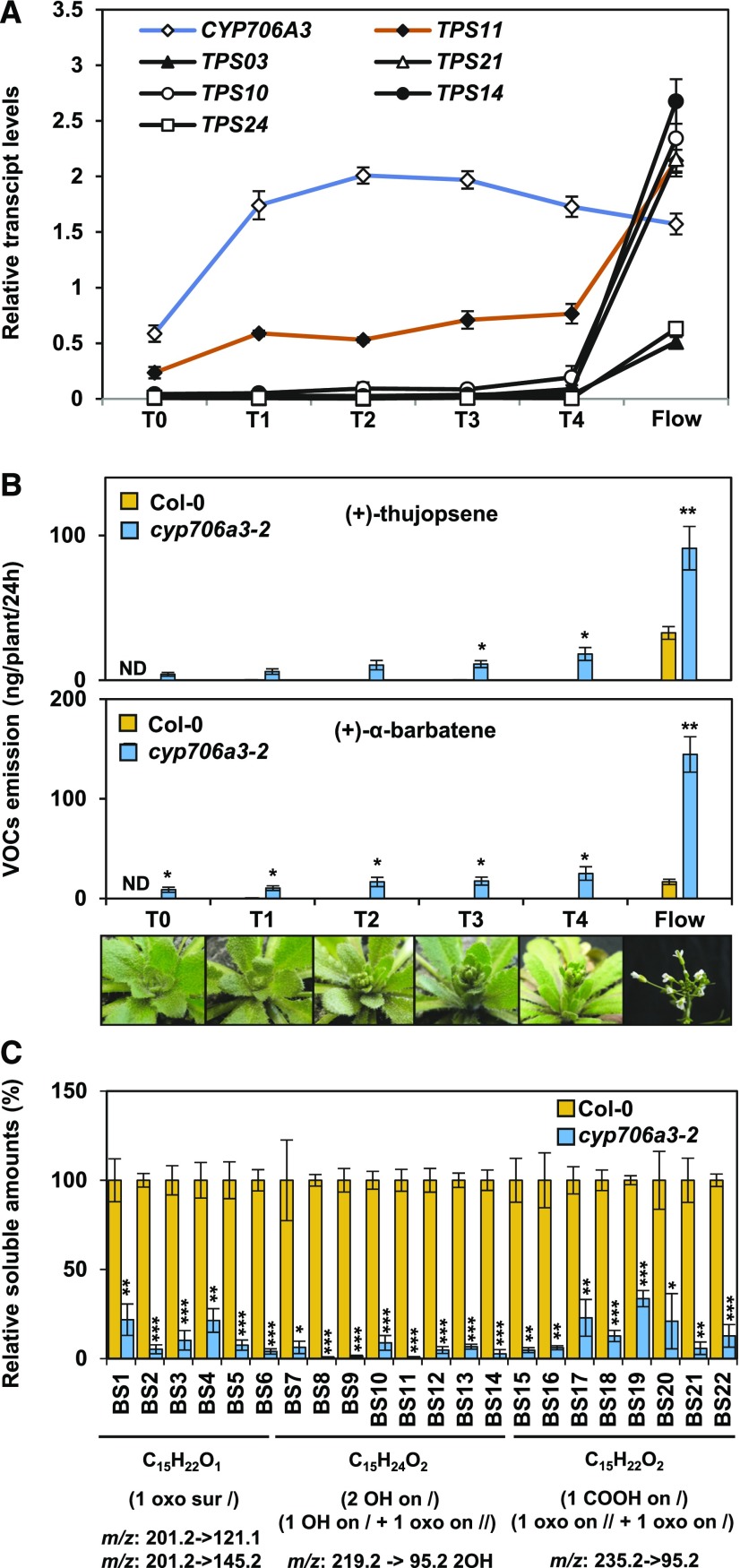
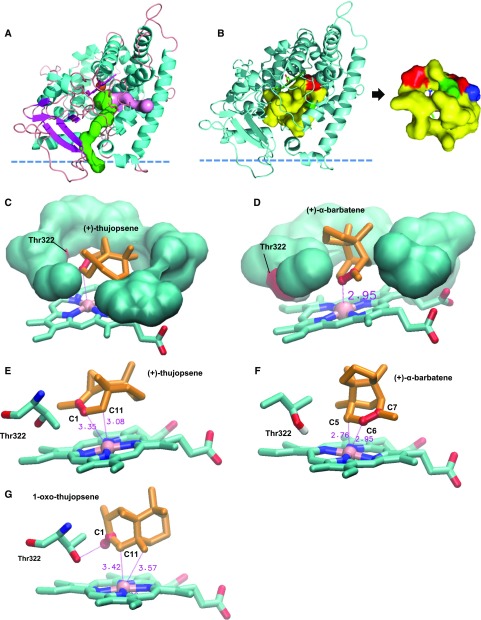
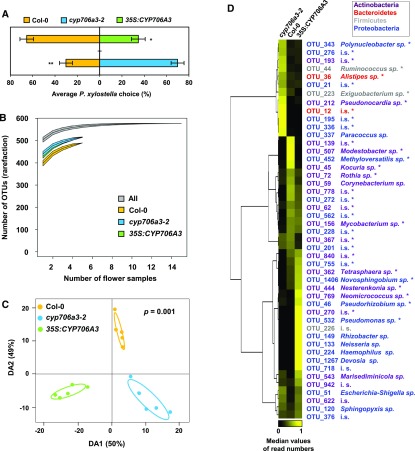
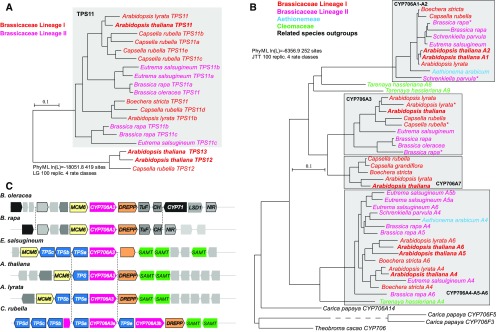
Flowers are essential but vulnerable plant organs, exposed to pollinators and florivores; however, flower chemical defenses are rarely investigated. We show here that two clustered terpene synthase and cytochrome P450 encoding genes (TPS11 and CYP706A3) on chromosome 5 of Arabidopsis (Arabidopsis thaliana) are tightly coexpressed in floral tissues, upon anthesis and during floral bud development. TPS11 was previously reported to generate a blend of sesquiterpenes. By heterologous coexpression of TPS11 and CYP706A3 in yeast (Saccharomyces cerevisiae) and Nicotiana benthamiana, we demonstrate that CYP706A3 is active on TPS11 products and also further oxidizes its own primary oxidation products. Analysis of headspace and soluble metabolites in cyp706a3 and 35S:CYP706A3 mutants indicate that CYP706A3-mediated metabolism largely suppresses sesquiterpene and most monoterpene emissions from opening flowers, and generates terpene oxides that are retained in floral tissues. In flower buds, the combined expression of TPS11 and CYP706A3 also suppresses volatile emissions and generates soluble sesquiterpene oxides. Florivory assays with the Brassicaceae specialist Plutella xylostella demonstrate that insect larvae avoid feeding on buds expressing CYP706A3 and accumulating terpene oxides. Composition of the floral microbiome appears also to be modulated by CYP706A3 expression. TPS11 and CYP706A3 simultaneously evolved within Brassicaceae and form the most versatile functional gene cluster described in higher plants so far.plantcell;31/12/2947/FX1F1fx1.
© 2019 American Society of Plant Biologists. All rights reserved.
Figures

Comment in
-
Attract or Defend: The CYP-Associated Versatility of Terpenoids.Plant Cell. 2019 Dec;31(12):2823. doi: 10.1105/tpc.19.00814. Epub 2019 Oct 18. Plant Cell. 2019. PMID: 31628165 Free PMC article. No abstract available.
References
-
- Abdalsamee M.K., Müller C. (2015). Uncovering different parameters influencing florivory in a specialist herbivore. Ecol. Entomol. 40: 258–268.
-
- Alonso J.M., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
