

Concatenation of 14-3-3 with partner phosphoproteins as a tool to study their interaction
- PMID: 31628352
- PMCID: PMC6802120
- DOI: 10.1038/s41598-019-50941-3
Concatenation of 14-3-3 with partner phosphoproteins as a tool to study their interaction
Abstract
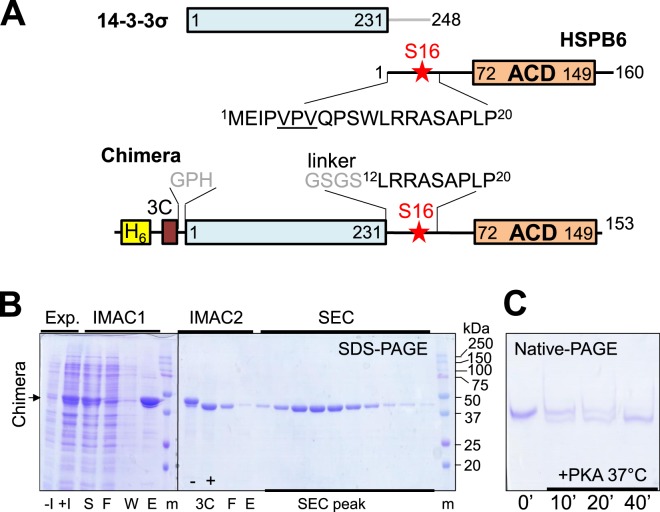
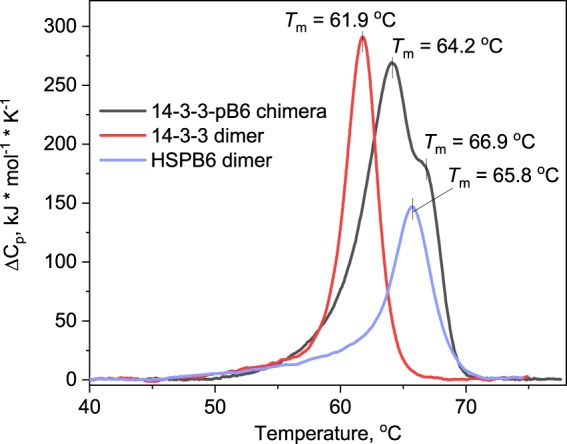
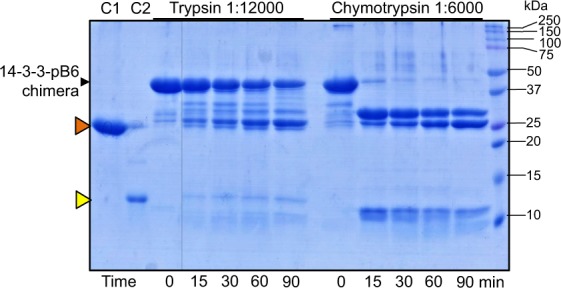
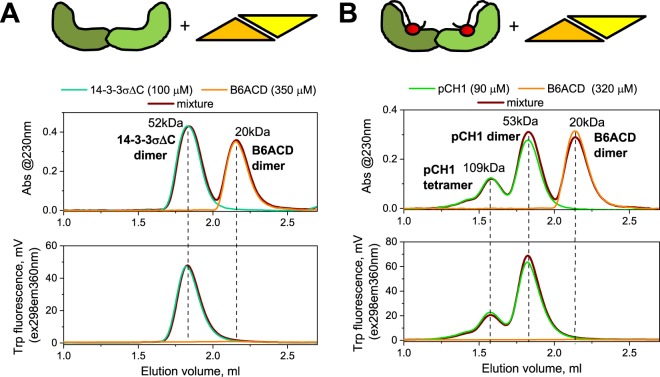
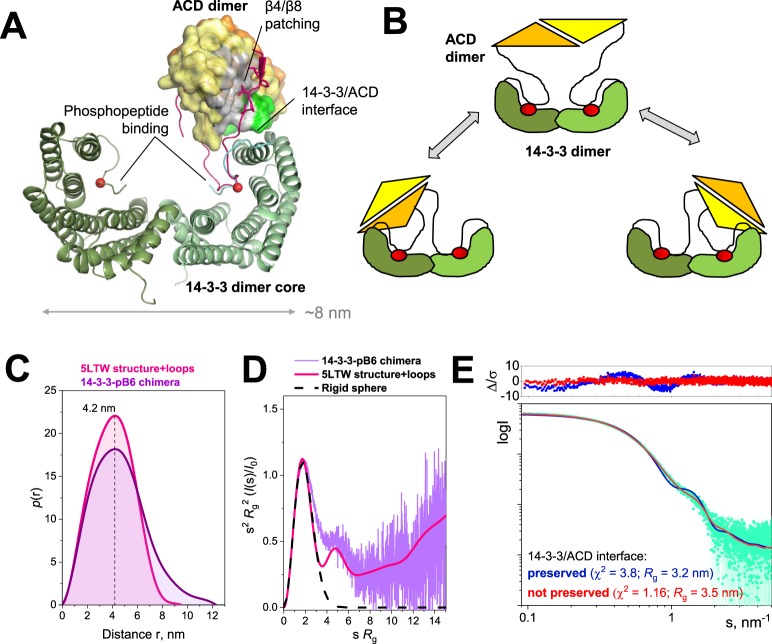
Regulatory 14-3-3 proteins interact with a plethora of phosphorylated partner proteins, however 14-3-3 complexes feature intrinsically disordered regions and often a transient type of interactions making structural studies difficult. Here we engineer and examine a chimera of human 14-3-3 tethered to a nearly complete partner HSPB6 which is phosphorylated by protein kinase A (PKA). HSPB6 includes a long disordered N-terminal domain (NTD), a phosphorylation motif around Ser16, and a core α-crystallin domain (ACD) responsible for dimerisation. The chosen design enables an unstrained binding of pSer16 in each 1433 subunit and secures the correct 2:2 stoichiometry. Differential scanning calorimetry, limited proteolysis and small-angle X-ray scattering (SAXS) support the proper folding of both the 14-3-3 and ACD dimers within the chimera, and indicate that the chimera retains the overall architecture of the native complex of 14-3-3 and phosphorylated HSPB6 that has recently been resolved using crystallography. At the same time, the SAXS data highlight the weakness of the secondary interface between the ACD dimer and the C-terminal lobe of 14-3-3 observed in the crystal structure. Applied to other 14-3-3 complexes, the chimeric approach may help probe the stability and specificity of secondary interfaces for targeting them with small molecules in the future.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
