

Differential functional patterns of memory CD4+ and CD8+ T-cells from volunteers immunized with Ty21a typhoid vaccine observed using a recombinant Escherichia coli system expressing S. Typhi proteins
- PMID: 31629569
- PMCID: PMC6994170
- DOI: 10.1016/j.vaccine.2019.10.020
Differential functional patterns of memory CD4+ and CD8+ T-cells from volunteers immunized with Ty21a typhoid vaccine observed using a recombinant Escherichia coli system expressing S. Typhi proteins
Abstract
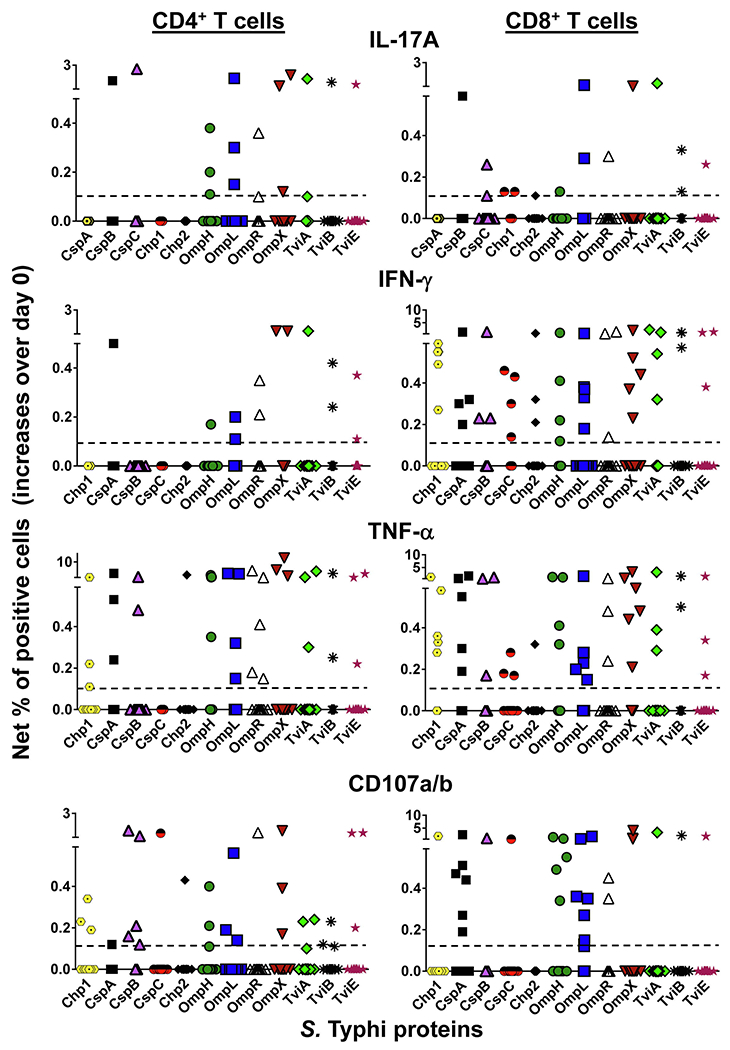
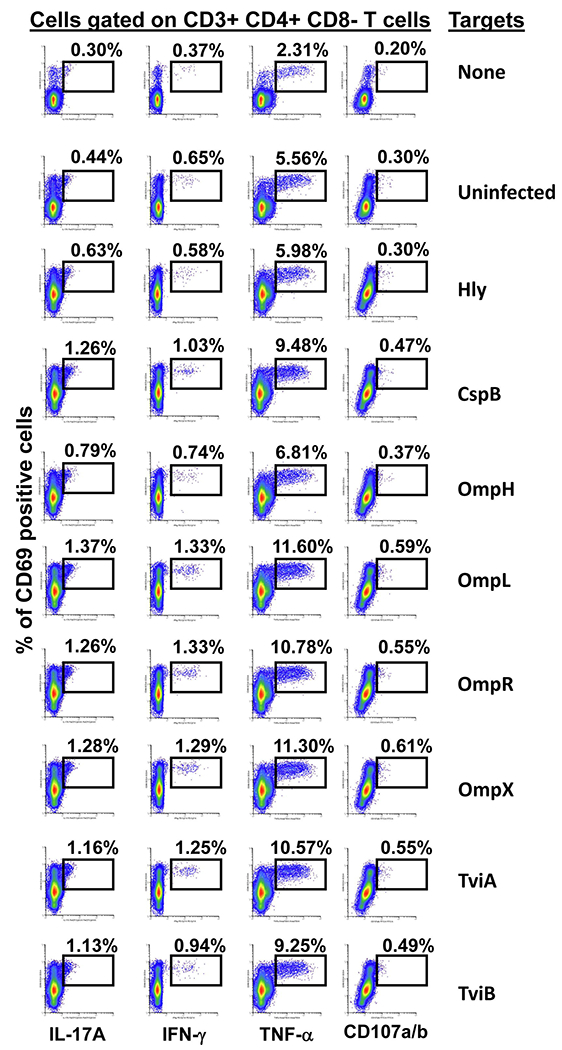
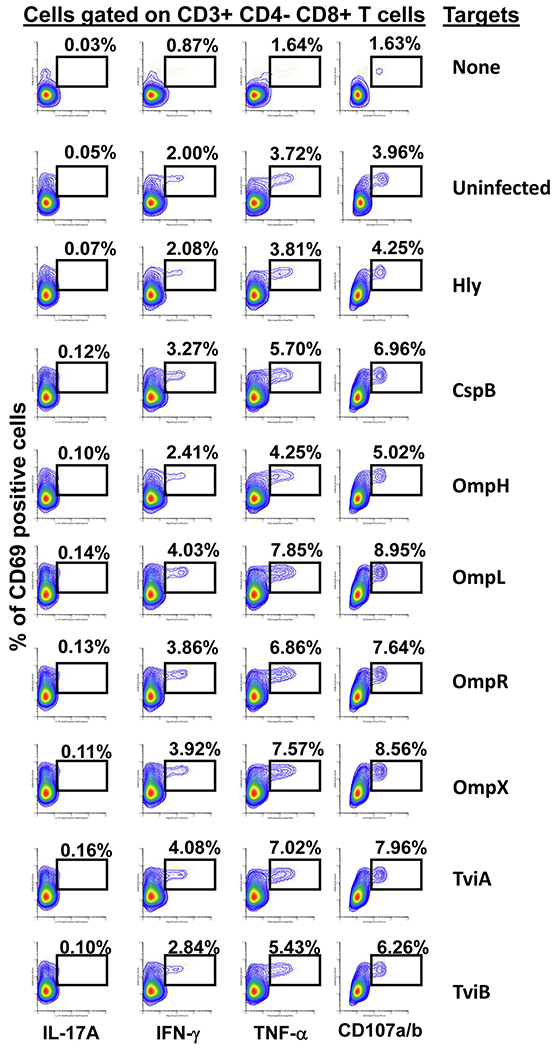
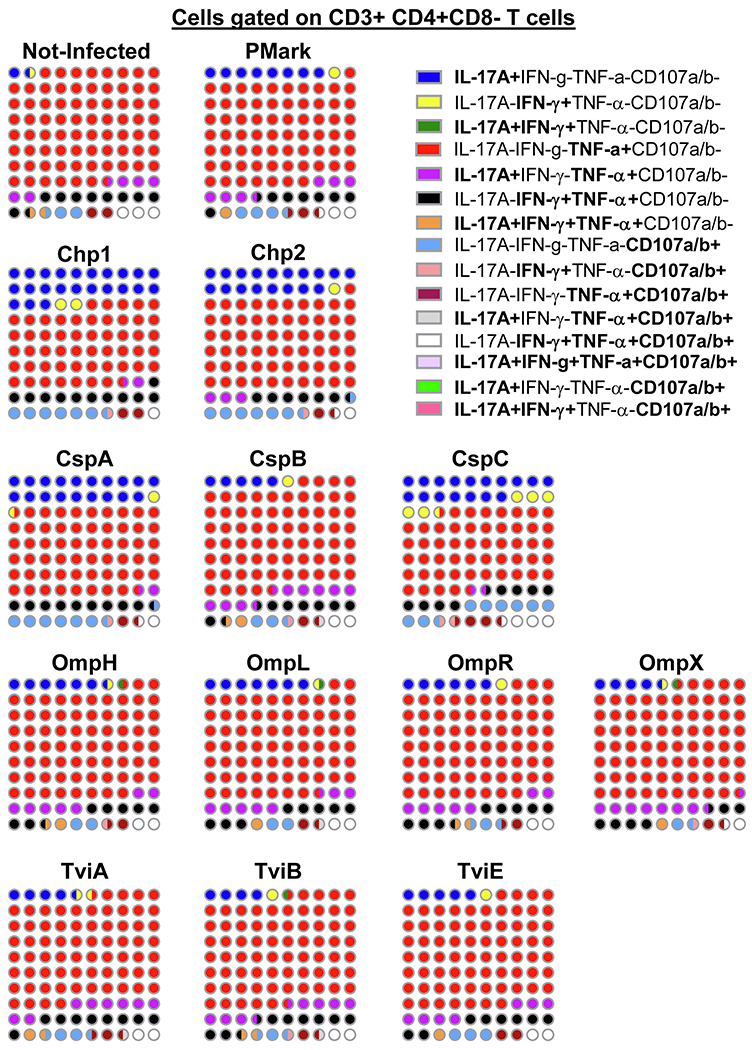
It is widely accepted that CD4+ and CD8+ T-cells play a significant role in protection against Salmonella enterica serovar Typhi (S. Typhi), the causative agent of the typhoid fever. However, the antigen specificity of these T-cells remains largely unknown. Previously, we demonstrated the feasibility of using a recombinant Escherichia coli (E. coli) expression system to uncover the antigen specificity of CD4+ and CD8+ T cells. Here, we expanded these studies to include the evaluation of 12 additional S. Typhi proteins: 4 outer membrane proteins (OmpH, OmpL, OmpR, OmpX), 3 Vi-polysaccharide biosynthesis proteins (TviA, TviB, TviE), 3 cold shock proteins (CspA, CspB, CspC), and 2 conserved hypothetical proteins (Chp 1 and Chp2), all selected based on the bioinformatic analyses of the content of putative T-cell epitopes. CD4+ and CD8+ T cells from 15 adult volunteers, obtained before and 42 days after immunization with oral live attenuated Ty21a vaccine, were assessed for their functionality (i.e., production of cytokines and cytotoxic expression markers in response to stimulation with selected antigens) as measured by flow cytometry. Although volunteers differed on their T-cell antigen specificity, we observed T-cell immune responses against all S. Typhi proteins evaluated. These responses included 9 proteins, OmpH, OmpR, TviA, TviE, CspA, CspB, CspC, Chp 1 and Chp 2, which have not been previously reported to elicit T-cell responses. Interestingly, we also observed that, regardless of the protein, the functional patterns of the memory T-cells were different between CD4+ and CD8+ T cells. In sum, these studies demonstrated the feasibility of using bioinformatic analysis and the E. coli expressing system described here to uncover novel immunogenic T-cell proteins that could serve as potential targets for the production of protein-based vaccines.
Keywords: Human; Recombinant E. coli; Salmonella; T-cells; Vaccine.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Conflict of interest
The authors declare no conflict of interest.
Declaration of interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures

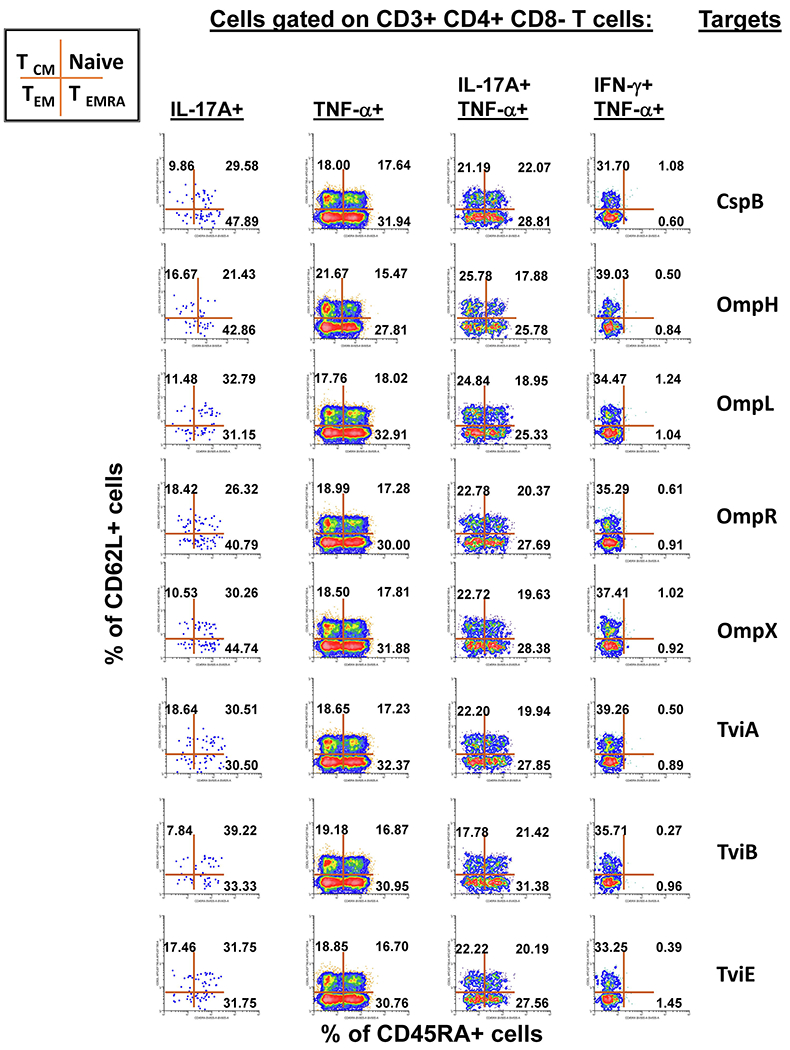
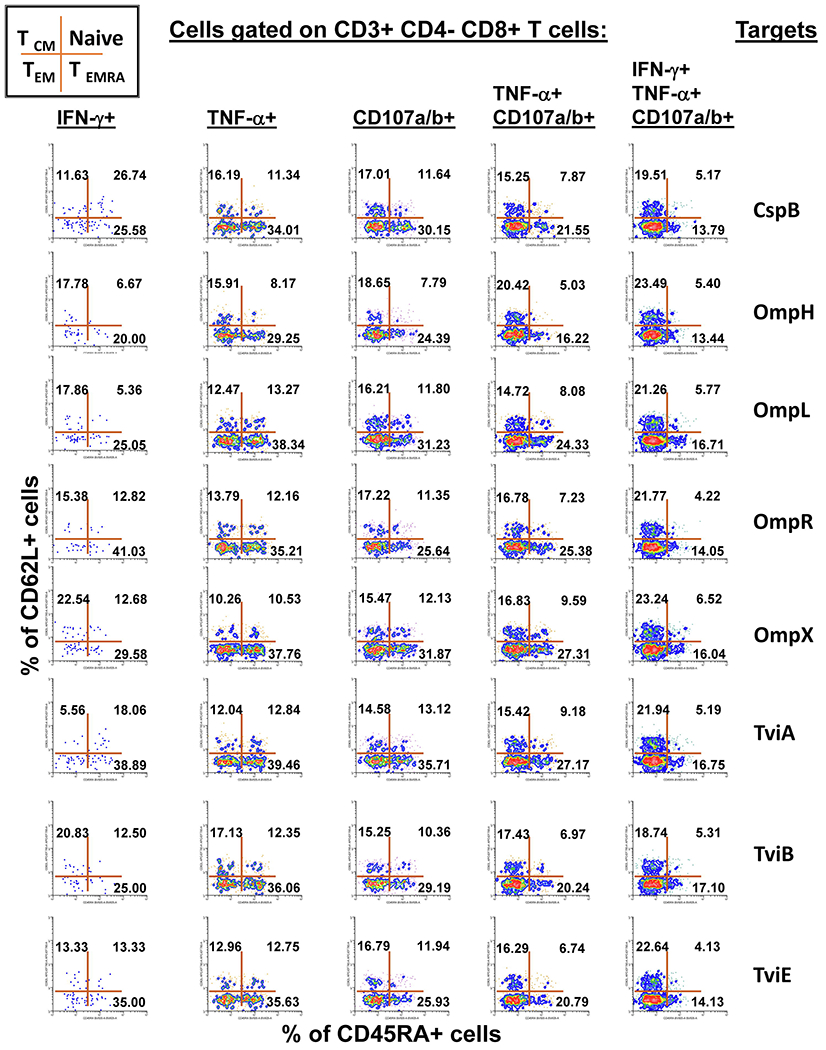
References
-
- Salerno-Goncalves R, Sztein MB. Cell-mediated immunity and the challenges for vaccine development. Trends in microbiology. 2006;14:536–42. - PubMed
-
- Sztein MB. Cell-mediated immunity and antibody responses elicited by attenuated Salmonella enterica Serovar Typhi strains used as live oral vaccines in humans. Clin Infect Dis. 2007;45 Suppl 1:S15–9. - PubMed
-
- Salerno-Goncalves R, Fernandez-Vina M, Lewinsohn DM, Sztein MB. Identification of a human HLA-E-restricted CD8+ T cell subset in volunteers immunized with Salmonella enterica serovar Typhi strain Ty21a typhoid vaccine. J Immunol. 2004;173:5852–62. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
