

Epigenome Regulation by Dynamic Nucleosome Unwrapping
- PMID: 31630896
- PMCID: PMC10168609
- DOI: 10.1016/j.tibs.2019.09.003
Epigenome Regulation by Dynamic Nucleosome Unwrapping
Abstract
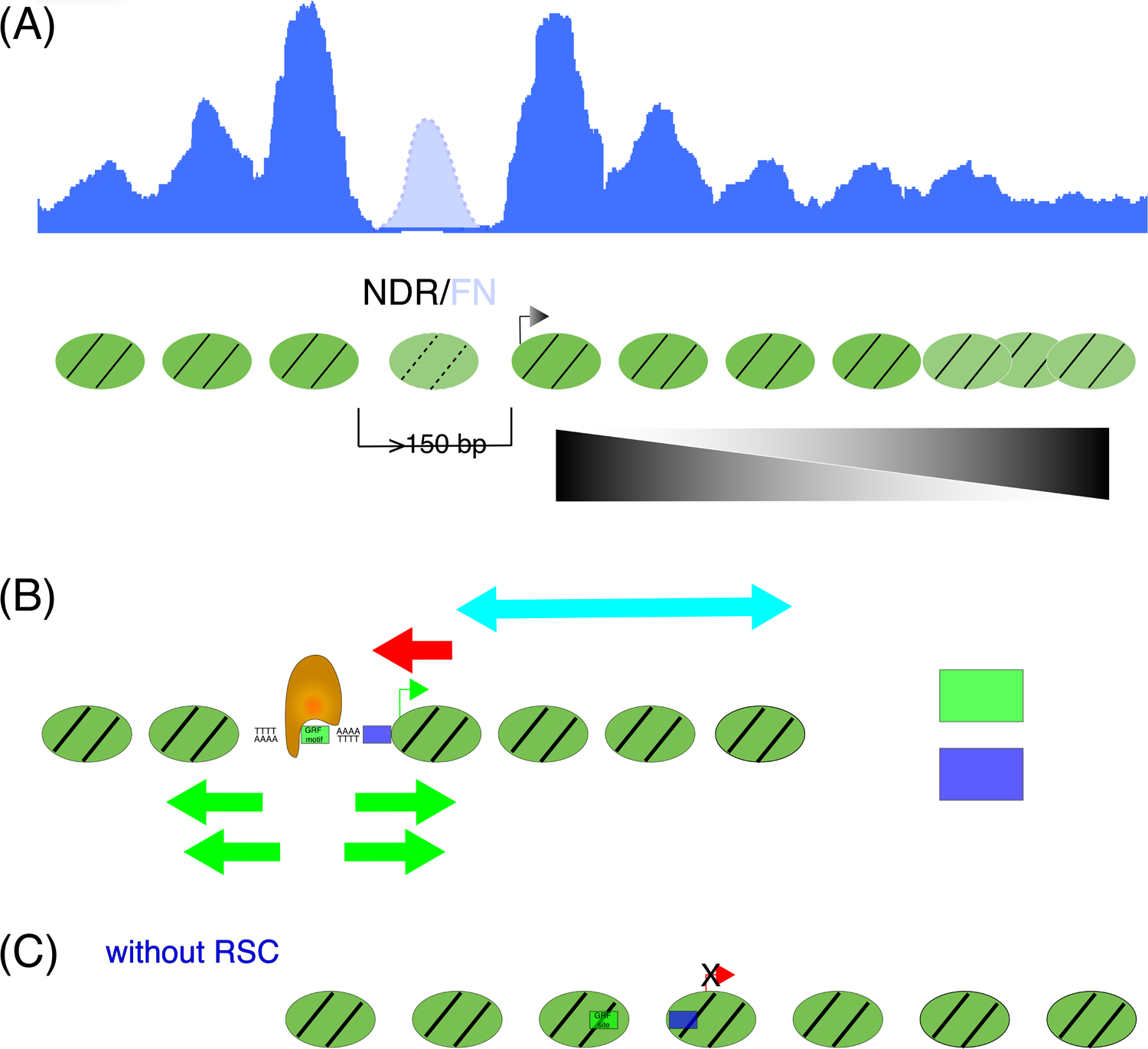
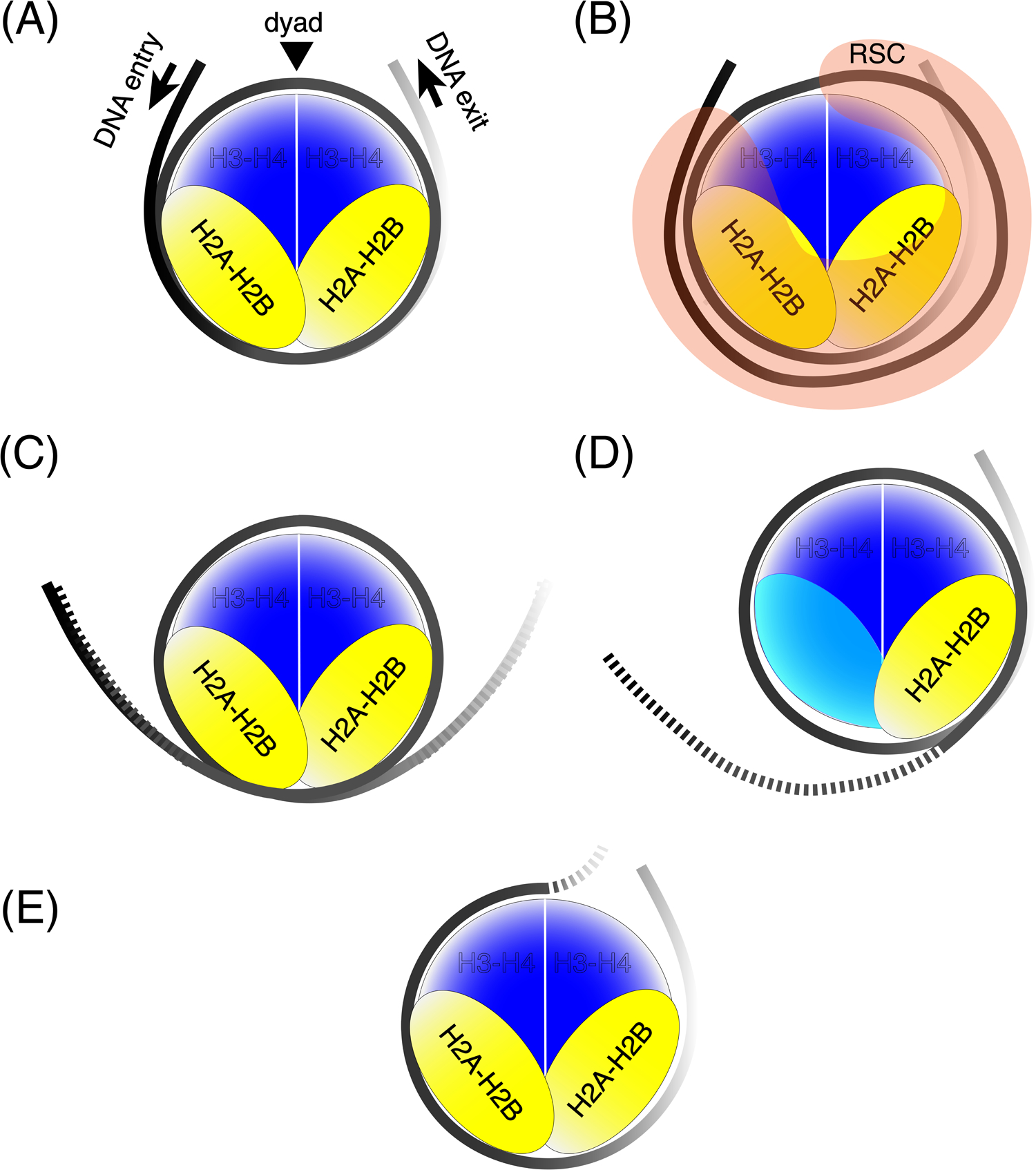
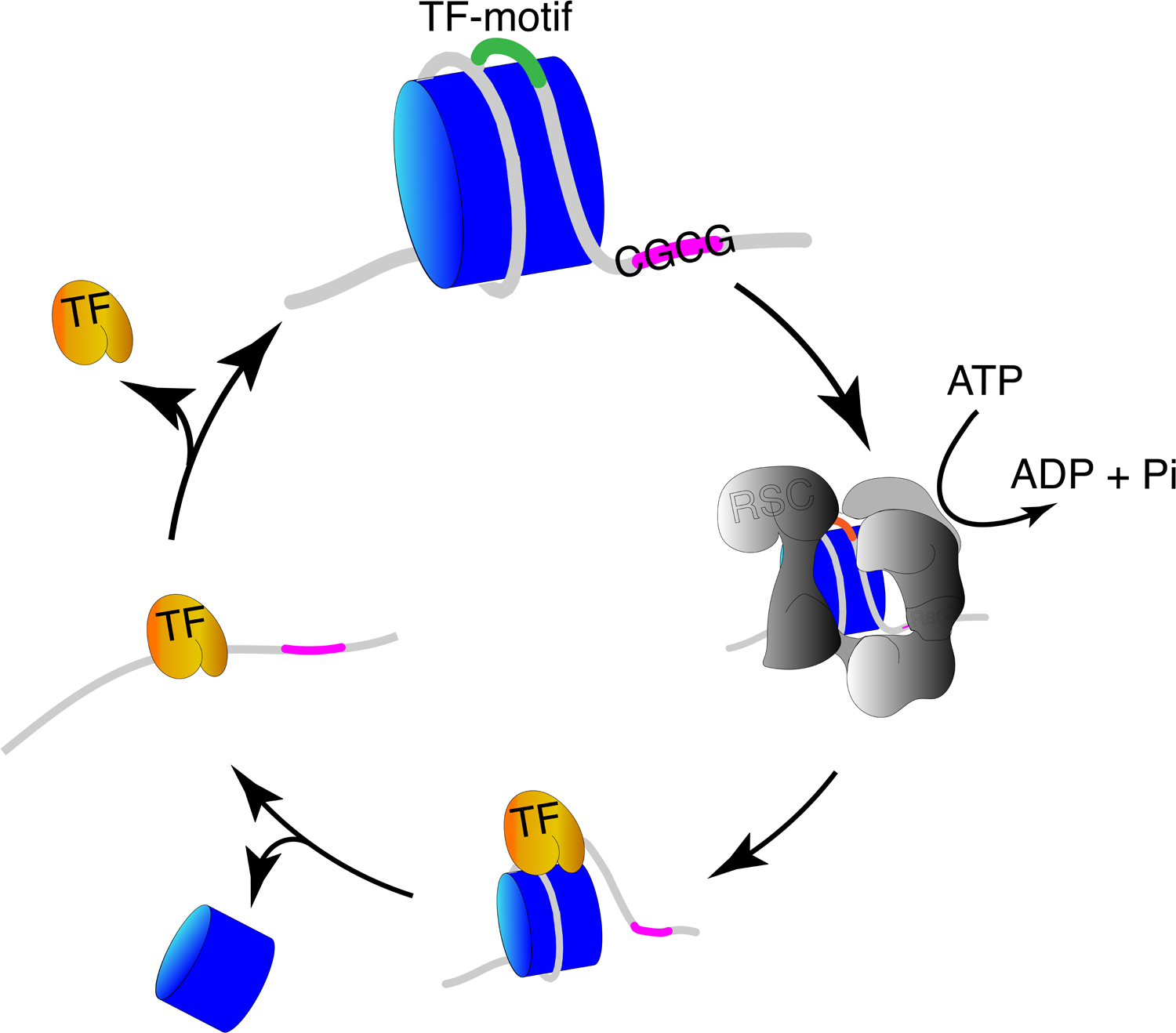
Gene regulation in eukaryotes requires the controlled access of sequence-specific transcription factors (TFs) to their sites in a chromatin landscape dominated by nucleosomes. Nucleosomes are refractory to TF binding, and often must be removed from regulatory regions. Recent genomic studies together with in vitro measurements suggest that the nucleosome barrier to TF binding is modulated by dynamic nucleosome unwrapping governed by ATP-dependent chromatin remodelers. Genome-wide occupancy and the regulation of subnucleosomal intermediates have gained recent attention with the application of high-resolution approaches for precision mapping of protein-DNA interactions. We summarize here recent findings on nucleosome substructures and TF binding dynamics, and highlight how unwrapped nucleosomal intermediates provide a novel signature of active chromatin.
Keywords: ATP-dependent remodeling; fragile nucleosome; nucleosome dynamics; structural epigenomics; transcription factors.
Copyright © 2019 Elsevier Ltd. All rights reserved.
Figures
References
-
- Davey CA et al. (2002) Solvent mediated interactions in the structure of the nucleosome core particle at 1.9 a resolution. J Mol Biol 319 (5), 1097–113. - PubMed
-
- Talbert PB et al. (2019) Old cogs, new tricks: the evolution of gene expression in a chromatin context. Nat Rev Genet 20 (5), 283–297. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
