

Terpene Synthases as Metabolic Gatekeepers in the Evolution of Plant Terpenoid Chemical Diversity
- PMID: 31632418
- PMCID: PMC6779861
- DOI: 10.3389/fpls.2019.01166
Terpene Synthases as Metabolic Gatekeepers in the Evolution of Plant Terpenoid Chemical Diversity
Abstract
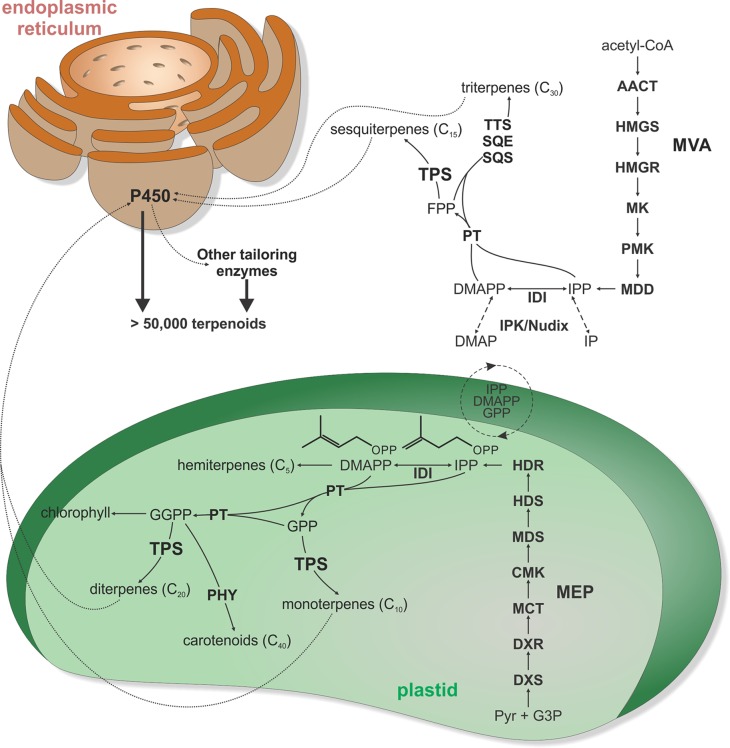
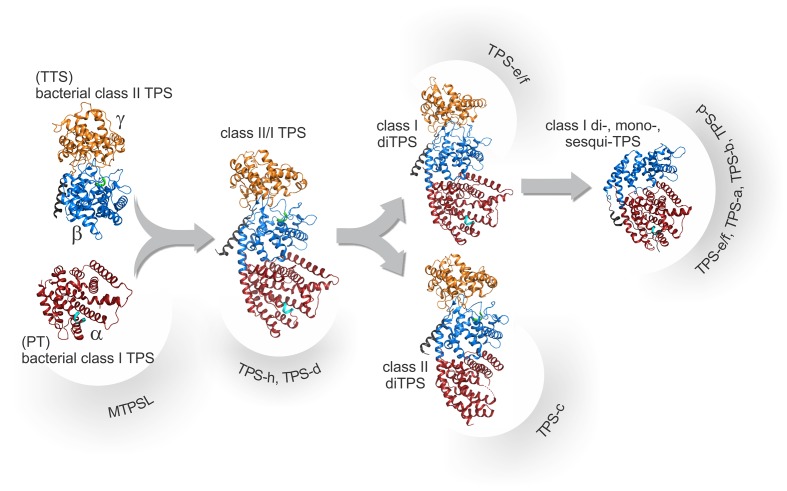
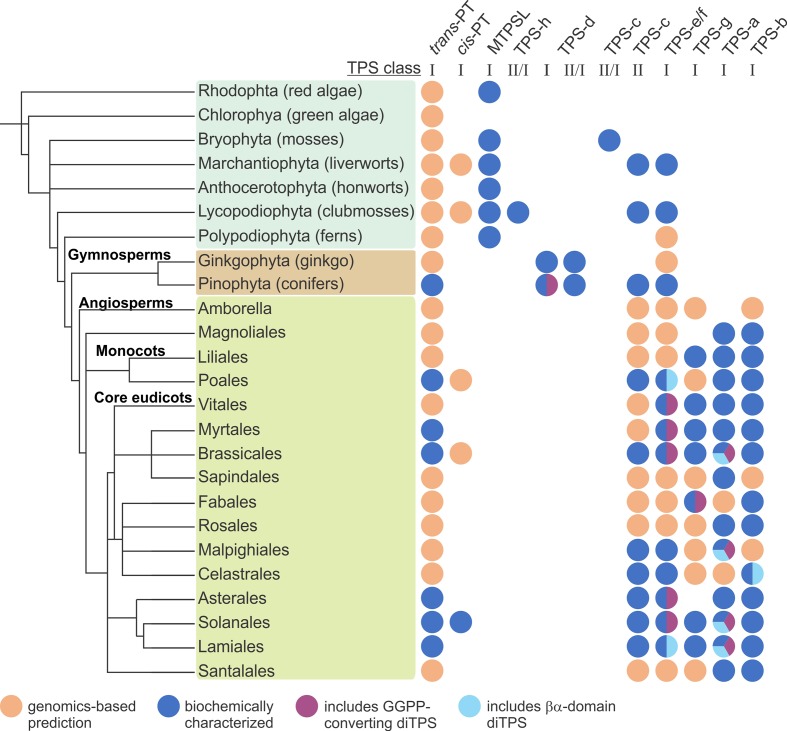
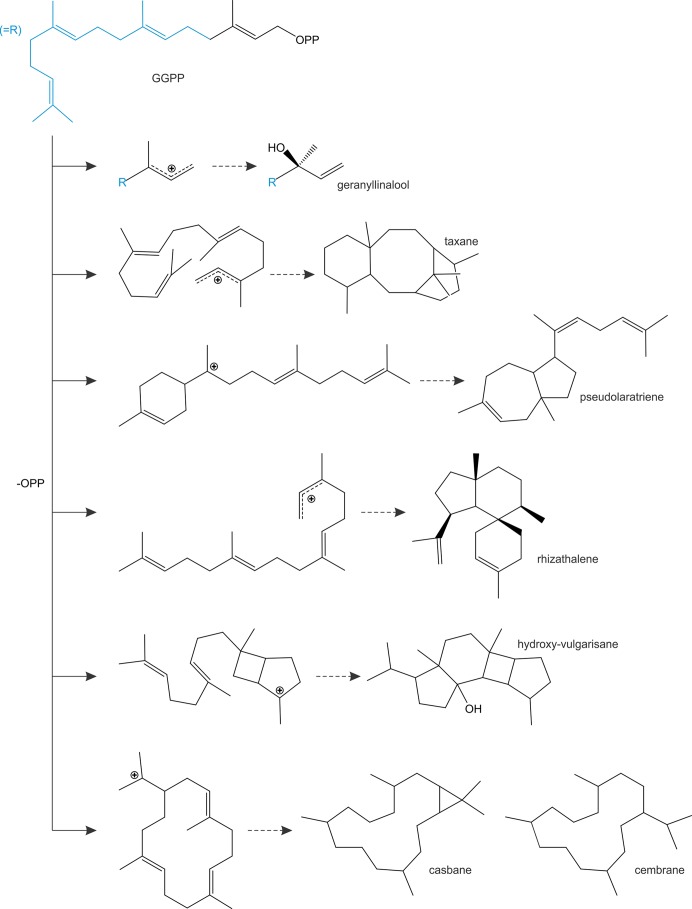
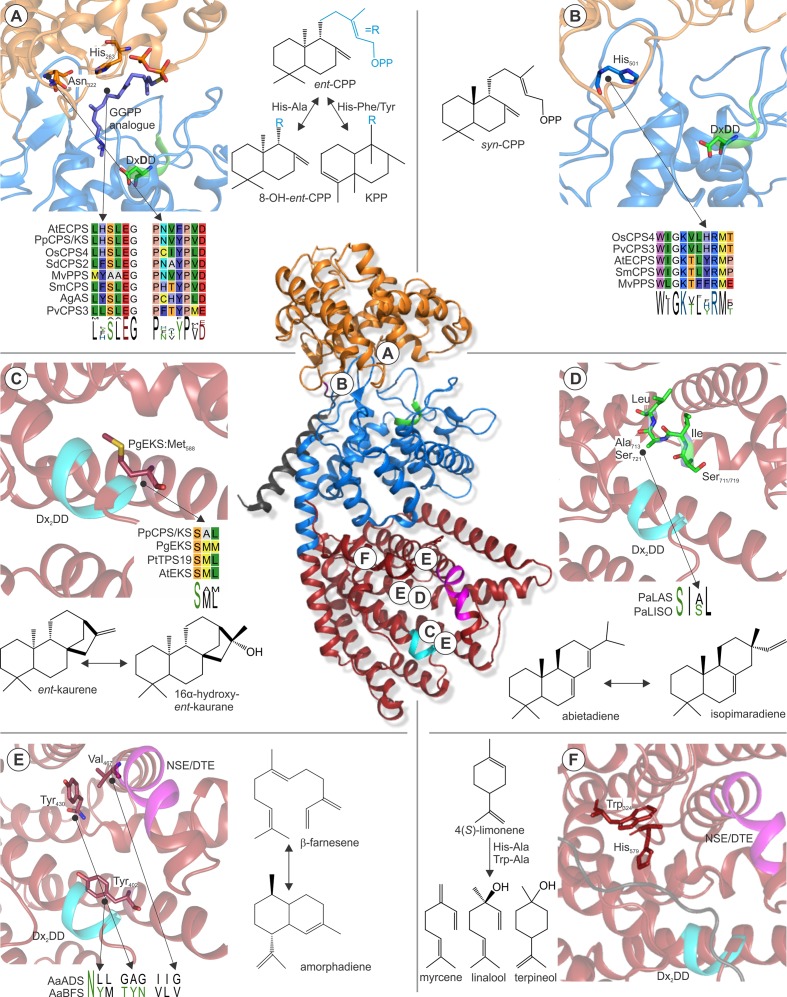
Terpenoids comprise tens of thousands of small molecule natural products that are widely distributed across all domains of life. Plants produce by far the largest array of terpenoids with various roles in development and chemical ecology. Driven by selective pressure to adapt to their specific ecological niche, individual species form only a fraction of the myriad plant terpenoids, typically representing unique metabolite blends. Terpene synthase (TPS) enzymes are the gatekeepers in generating terpenoid diversity by catalyzing complex carbocation-driven cyclization, rearrangement, and elimination reactions that enable the transformation of a few acyclic prenyl diphosphate substrates into a vast chemical library of hydrocarbon and, for a few enzymes, oxygenated terpene scaffolds. The seven currently defined clades (a-h) forming the plant TPS family evolved from ancestral triterpene synthase- and prenyl transferase-type enzymes through repeated events of gene duplication and subsequent loss, gain, or fusion of protein domains and further functional diversification. Lineage-specific expansion of these TPS clades led to variable family sizes that may range from a single TPS gene to families of more than 100 members that may further function as part of modular metabolic networks to maximize the number of possible products. Accompanying gene family expansion, the TPS family shows a profound functional plasticity, where minor active site alterations can dramatically impact product outcome, thus enabling the emergence of new functions with minimal investment in evolving new enzymes. This article reviews current knowledge on the functional diversity and molecular evolution of the plant TPS family that underlies the chemical diversity of bioactive terpenoids across the plant kingdom.
Keywords: natural products; plant chemical diversity; plant specialized metabolism; terpene synthases; terpenoid biosynthesis; terpenoids.
Copyright © 2019 Karunanithi and Zerbe.
Figures

References
-
- Andersen-Ranberg J., Kongstad K. T., Nielsen M. T., Jensen N. B., Pateraki I., Bach S. S., et al. (2016). Expanding the landscape of diterpene structural diversity through stereochemically controlled combinatorial biosynthesis. Angew. Chem. Int. Ed. Engl. 55, 2142–2146. 10.1002/anie.201510650 - DOI - PMC - PubMed