

Protein aggregation in bacteria
- PMID: 31633151
- PMCID: PMC7053576
- DOI: 10.1093/femsre/fuz026
Protein aggregation in bacteria
Abstract
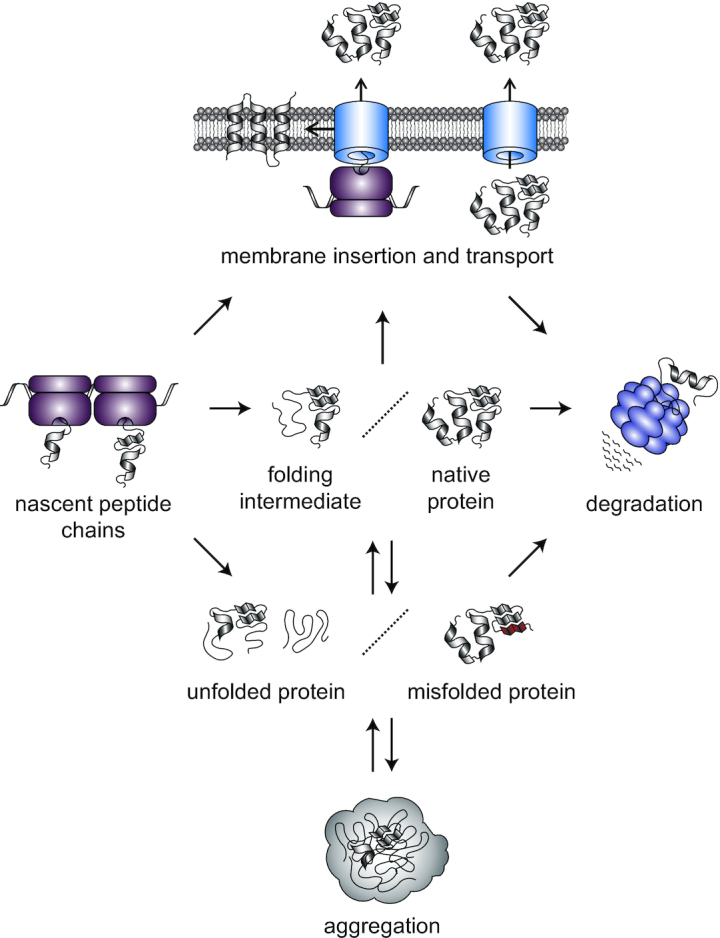
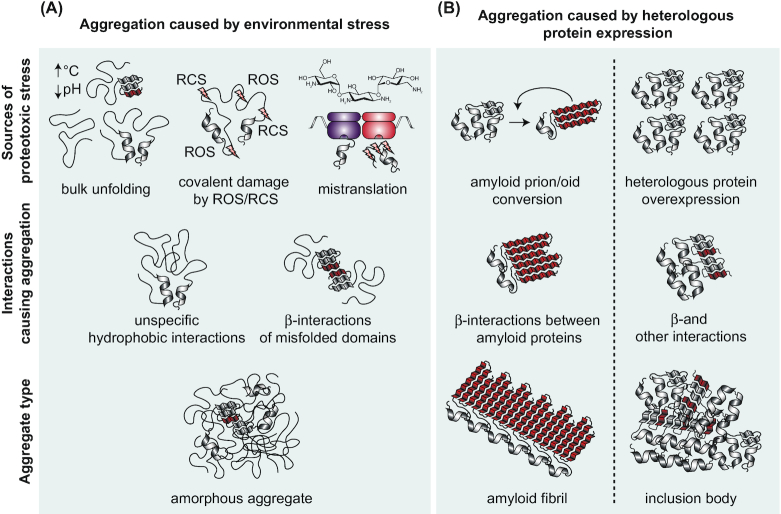
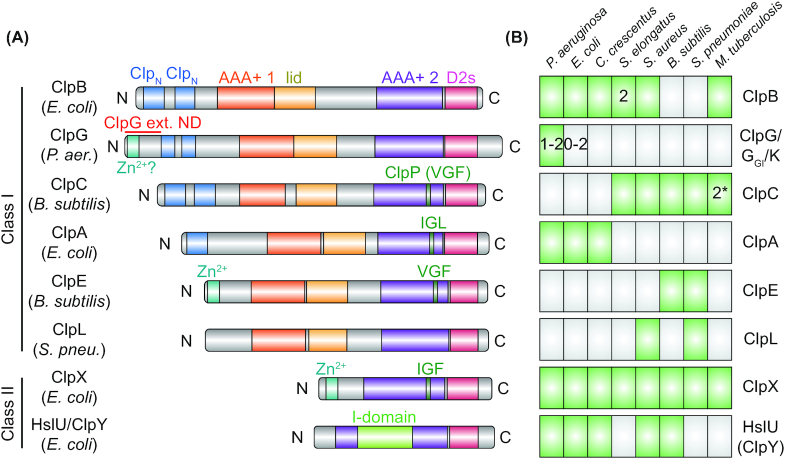
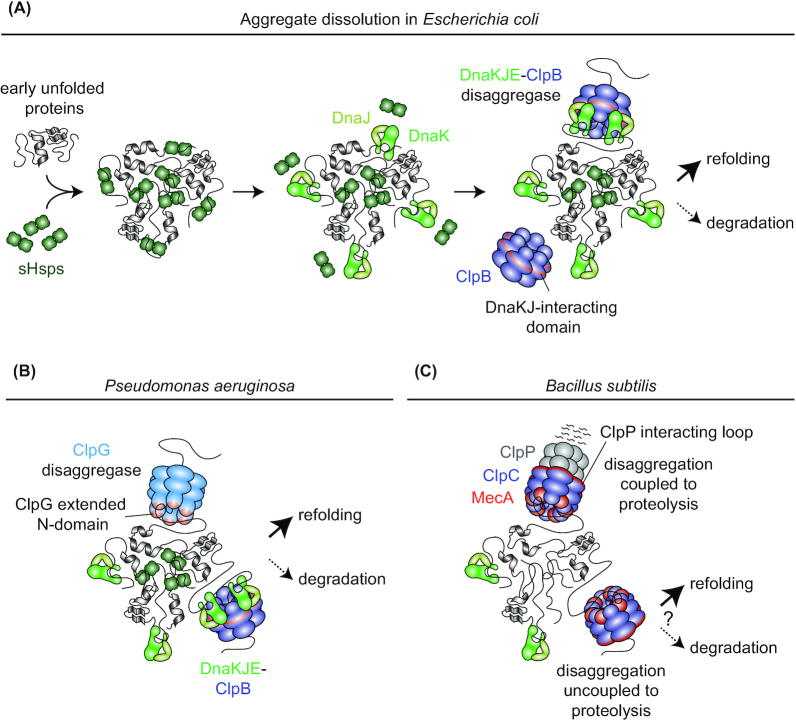
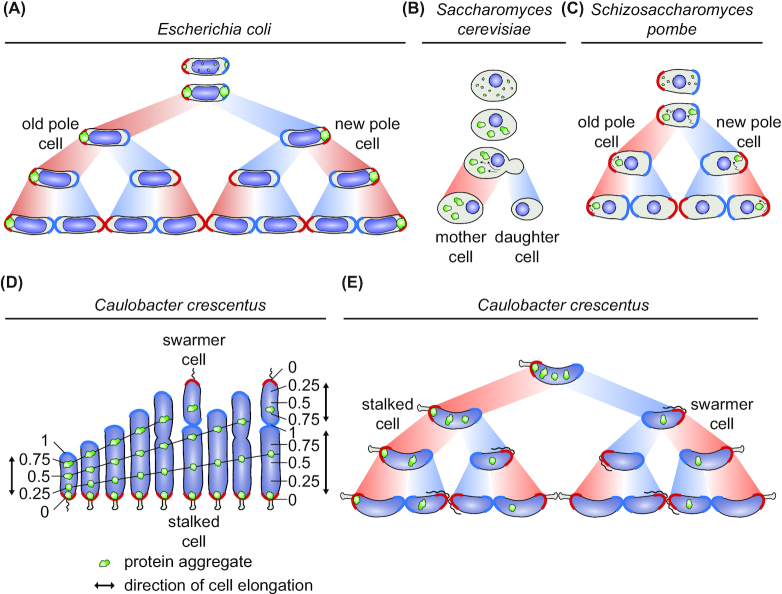
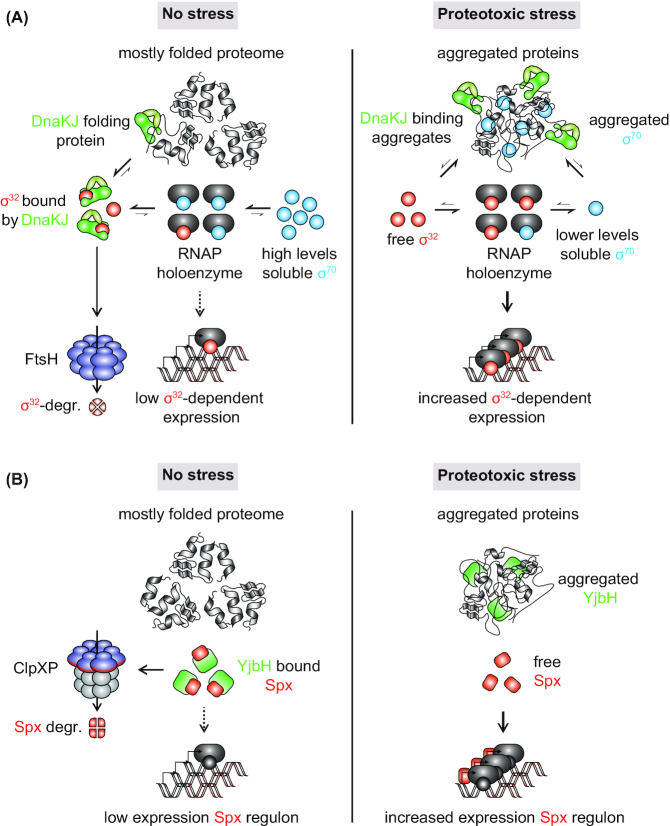
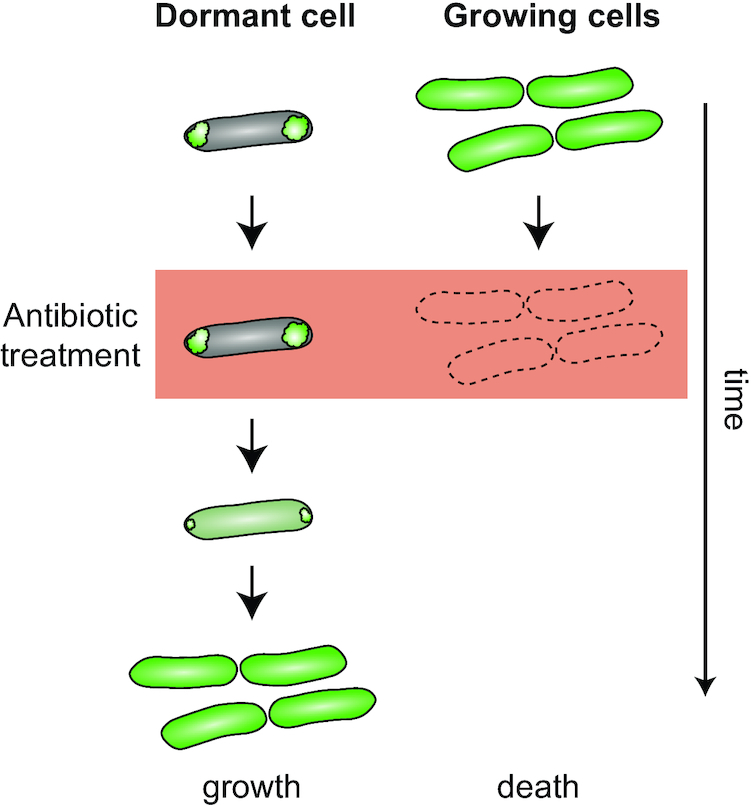
Protein aggregation occurs as a consequence of perturbations in protein homeostasis that can be triggered by environmental and cellular stresses. The accumulation of protein aggregates has been associated with aging and other pathologies in eukaryotes, and in bacteria with changes in growth rate, stress resistance and virulence. Numerous past studies, mostly performed in Escherichia coli, have led to a detailed understanding of the functions of the bacterial protein quality control machinery in preventing and reversing protein aggregation. However, more recent research points toward unexpected diversity in how phylogenetically different bacteria utilize components of this machinery to cope with protein aggregation. Furthermore, how persistent protein aggregates localize and are passed on to progeny during cell division and how their presence impacts reproduction and the fitness of bacterial populations remains a controversial field of research. Finally, although protein aggregation is generally seen as a symptom of stress, recent work suggests that aggregation of specific proteins under certain conditions can regulate gene expression and cellular resource allocation. This review discusses recent advances in understanding the consequences of protein aggregation and how this process is dealt with in bacteria, with focus on highlighting the differences and similarities observed between phylogenetically different groups of bacteria.
Keywords: aggregate inheritance; cellular aging; disaggregases; molecular chaperones; protein aggregation; stress adaptation.
© FEMS 2019.
Figures
References
-
- Acebrón SP, Martín I, del Castillo Uet al.. DnaK-mediated association of ClpB to protein aggregates. A bichaperone network at the aggregate surface. FEBS Lett. 2009;583:2991–6. - PubMed
-
- Ackermann M, Stephen CS, Jenal U. Senescence in a bacterium with asymmetric division. Science. 2003;300:1920. - PubMed
-
- Aguzzi A, Lakkaraju AKK, Frontzek K. Toward therapy of human prion diseases. Annu Rev Pharmacol Toxicol. 2018;58:331–51. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
